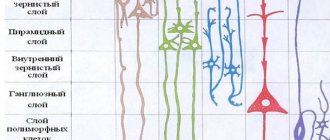
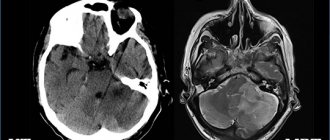
Характерная особенность человеческого мозга – невероятный размер коры и сложная складчатость. Кора – наиболее развитая область головного мозга, отвечающая за нерефлекторную деятельность (память, восприятие, познание, мышление и т. п.).
Формирование корково-подкорковых структур происходит во время эмбрионального развития, обеспечивая возможность размещения коры в ограниченном объеме черепной коробки. Извилины (гири) и борозды (сульчи) составляют её сложенную поверхность. Патологические изменения в размерах или складках коры приводят к тяжелой умственной неполноценности и трудноизлечимой эпилепсии. Следовательно, корковое расширение и складчатость рассматриваются как ключевые процессы в эволюции мозга.
Борозды и извилины: формирование и функции
Борозды и извилины в нейроанатомии, придающие головному мозгу морщинистый вид, служат двум важнейшим функциям. Они помогают увеличить площадь поверхности коры, что позволяет большему количеству нейронам уплотняться в неё и усиливать способность мозга обрабатывать информацию. Борозды и извилины головного мозга образуют деления, создавая границы между долями мозга, разделяя его на два полушария.
Основные борозды:
- Межполушарная щель – глубокая борозда по центру мозга, в которой находится мозолистое тело.
- Сильвиевая щель (латеральная борозда) разделяет теменную и лобную доли.
- Роландова щель (центральная борозда), отделяющая веретенообразную извилину и извилину гиппокампа на нижней поверхности височных долей.
- Теменно-затылочная — разделяет теменную и затылочную доли.
- Шпорная щель (шпороподобная борозда или выделяющаяся фиссура) – расположена в затылочных долях, делит зрительную кору.
Основные извилины мозга:
- Ангулярная извилина теменной доли помогает в обработке слухового и визуального распознавания.
- Извилина Брока (центр Брока) – область мозга, расположенная в левой лобной доле у большинства людей, которая контролирует функции, связанные с воспроизведением речи.
- Поясная извилина – арочная складка, расположенная над мозолистым телом, является компонентом лимбической системы и обрабатывает сенсорный ввод относительно эмоций, регулирует агрессивное поведение.
- Веретенообразная извилина находится в височной и затылочной долях и состоит из боковых и медиальных частей. Считается, что играет роль в распознавании слов и лиц.
- Гиппокампальная извилина складывается на внутренней поверхности височной доли, которая граничит с гиппокампом. Играет важную роль для памяти.
- Язычная извилина в затылочной доле, участвующая в зрительной обработке. Она ограничена коллатеральной бороздой и шпорной щелью. Спереди соприкасается с парарпопампальной извилиной, и вместе они образуют медиальную часть веретенообразной извилины.
По мере развития эмбриона извилины и борозды формируются с появлением углублений на поверхности коры. Не все извилины развиваются одновременно. Первичная форма образуется, начиная с 10 недели беременности (у человека), затем развиваются вторичные и третичные. Наиболее выдающаяся борозда – латеральная. За ней следует центральная, отделяющая моторную кору (прецентральную извилину) от соматосенсорной коры (постцентральной извилины). Большинство кортикальных борозд и извилин головного мозга, анатомия которых начинает складываться между 24 и 38 неделями беременности, продолжают расти и развиваться после того, как новорожденный появится на свет.
Раннее состояние головного мозга оказывает сильное влияние на конечный уровень гирификации. В частности, существует обратная связь между корковой толщиной и гирификацией. Участки мозга с низким значением толщины имеют более высокий уровень гирификации. Также верно и обратное, что участки мозга с высоким значением толщины (например, утолщение коры гиппокампальных извилин головного мозга) – низкий уровень гирификации.
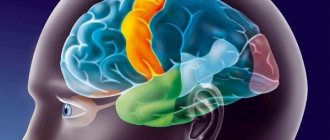
Доли мозга и их функции
Каждое из полушарий разделено на четыре доли: лобную, теменную, височную и затылочную. Большинство функций мозга полагаются на разные области по всему мозгу, которые работают вместе, но каждая доля выполняется основную часть относительно определенных функций.
Лобная доля расположена в самой передней области коры головного мозга, отделена от теменной доли центральной бороздой, от височной – боковой. В области, как правило, сосредоточены наиболее значимые для человека исполнительные функции, включая регулирование эмоций, планирование, рассуждение, решение проблем.
Теменная доля ответственна за интеграцию сенсорной информации, включая контакт, температуру, давление, боль. Из-за обработки, возникающей в париетальной доле, возможно различать прикосновение двух объектов в близлежащих точках (а не как один объект). Этот процесс называют двухточечным.
Височная доля также содержит области, участвующие в обработке сенсорной информации, особенно важные для слуха, распознавания языка, формирования воспоминаний. Первичная слуховая кора получает аудио информацию через уши и вторичные области и обрабатывает данные так, чтобы человек понимал то, что он слышит (слова, смех, плач и так далее). Медиальная (ближе к центру мозга) часть содержит гиппокамп – зону, важную для памяти, обучения, восприятия эмоций. Некоторые области височной доли обрабатывают сложную визуальную информацию, включая лица и сцены.
Значение, роль коры больших полушарий головного мозга человека
В статье мы рассмотрим локализацию функций, участки, анализаторы, поля, участки,
области зоны коры больших полушарий головного мозга человека (мужчины, женщины). Неврологи, невропатологи, рефлексотерапевты, рефлексологи выделяют 4 основных положения, применительно к практической деятельности невропатолога, современного учения о локализации функций в коре головного мозга.
1. Очень сложная морфологическая и функциональная дифференциация коры больших полушарий головного мозга. Лобная доля больше отвечает за двигательные функции. Теменная, затылочная и височная зоны больше отвечают за чувствительные функции.
2. Динамичность и относительность локализаций функций коры головного мозга. Определенный участок коры головного мозга, обеспечивая какую-то одну функцию, в то же время в разнообразных сочетаниях с другими ее полями может участвовать в осуществлении различных корковых функций и образовывать новые кортикальные связи. Это имеет значение в процессах компенсации при таких состояниях, как поражение коры головного мозга, нарушение коры головного мозга, смерть или повреждение коры головного мозга, отмирание, незрелость коры головного мозга.
3. Формирование специальных корковых областей в процессе практической деятельности.
Клеточные механизмы, приводящие к расширению и складыванию коры головного мозга
Строение мозга человека отличает его от прочих млекопитающих, и по этой причине может объяснять его уникальные умственные способности по сравнению с другими животными. Количество складок в коре, возможно, коррелирует с некоторыми специфическими когнитивными, сенсорными, двигательными способностями. Хотя нет четкого объяснения того, каким образом происходит уникальное разделение человеческого мозга на борозды и извилины. Сегодня имеется прогресс в понимании чрезвычайно сложных процессов в мозге, кора которого строится с таким количеством борозд и извилин. Несмотря на то, что у всех клеток одна и та же ДНК, образуются разные нервные стволовые клетки. Именно их работа с различными свойствами создает основную структуру мозга, состоящую из нейронов и глиальных клеток.
Теленцефалический нейроэпителий
Рост мозга происходит посредством двух видов стволовых клеток – нейронных стволовых клеток и нейронных предшественников. Обе эти формы образуют нейроны, которые становятся постоянными в мозге, а также промежуточные клетки, создающие строительный материал для построения мозга. Четыре различных типа стволовых клеток определяют строение коры.
В период раннего эмбрионального развития расширение рострального домена нервной трубки приводит к появлению двух телэнцефальных пузырей. Дорзальная половина этих пузырей молекулярно определяется как зачаток коры головного мозга. На этом этапе кортикальный зачаток состоит исключительно из монослоя нейроэпителиальных клеток-предшественников. Они сильно поляризованы и прикреплены друг к другу плотными соединениями на уровне апикального домена (внутренней поверхности телэнцефалического пузыря) и перемещают клеточное ядро между апикальной (верхушечной) и базальной (нижней) стороной нейроэпителия в согласованности с клеточным циклом.
- базально-направленное движение во время G1-фазы;
- базальное положение во время S-фазы;
- апикально-направленное движение во время G2-фазы;
- митоз на апикальной поверхности.
Циклическое движение известно как межкинетическая ядерная миграция и полностью асинхронно между нейроэпителиальными клетками, придавая нейроэпителию псевдостратированный вид. Клетки подвергаются только симметричным самоагрессирующим делениям, при этом каждое деление генерирует две дочерние клетки, следовательно, экспоненциально увеличивая их число. Поскольку они являются основополагающими клетками-предшественниками коры головного мозга, размер их объединения определяет количество производных нейрогенных клеток-предшественников и конечное число кортикальных нейронов, и, следовательно, он оказывает фундаментальное влияние на размер зрелой коры головного мозга. Увеличение количества приводит к расширению площади поверхности и формированию нейроэпителия.
Распространение и нейрогенез
Непосредственно перед началом нейрогенеза нейроэпителиальные клетки-предшественники начинают терять плотные соединения, и приобретать признаки, типичные для глиальных клеток (включая экспрессию липид-связывающего белка мозга, виментина и Pax6), становясь, таким образом, апикальными радиальными глиальными клетками (АРГК). Они также подвергаются межкинетической ядерной миграции, делятся на верхушечной поверхности развивающейся коры и на этом раннем этапе также подвергаются самоусиливающимся делениям.
Однако постепенно они начинают делиться асимметрично, чтобы генерировать одну себе подобную клетку плюс другую клетку. Эти новые клетки накапливаются в базальной части коркового зачатка, тогда как клеточные тела АРГК остаются на апикальной стороне, образуя желудочковую зону (ЖЗ). С накоплением клеток выше ЖЗ процесс АРГК продлевается, оставаясь присоединенным к базальной пластине, и теперь называется радиальной глией. Асимметричные АРГК-деления генерируют один АРГК плюс один нейрон или одну промежуточную клетку-предшественник. Промежуточные клетки-предшественники (вторичные клетки-предшественники без апикально-базальной полярности) не подвергаются межкинетической ядерной миграции, делятся в слое, расположенном в области желудочковой зоны, субвентрикулярной зоны (СВЗ), и все они выражают транскрипционный фактор (Tbr2).
Однако из-за того, что каждый нейрон сам потребляет при митозе, их относительное количество по сравнению с АРГК достаточно низкое. Промежуточные клетки-предшественники в коре головного мозга генерируют большинство кортикальных возбуждающих нейронов. По мере развития нейрогенеза потребность в расширении/обновлении АРГК снижается, и повышается в производстве нейронов. Дополнительно к расширенной желудочковой зоне утолщается субвентрикулярная зона, населенная в изобилии базальными предшественниками, особенно на поздних стадиях нейрогенеза. В результате происходит расщепление СВЗ на внутреннюю и внешнюю часть. Внешняя часть содержит большое разнообразие типов клеток-предшественников с высоким потенциалом развития, что является ключевым фактором для расширения и формирования коры.
Обсуждение
Несмотря на практическую важность (до 25% всех низкозлокачественных и до 10% всех высокозлокачественных глиом расположены в островковой доле [10]) и функциональную сложность (островковая доля окружена расположенными вокруг сильвиевой щели центрами речи Брока и Вернике, первичной моторной и сенсорной корой зоны лица, а также проводящими путями, соединяющими данные области) островка, в настоящее время доступны лишь несколько публикаций, посвященных изучению анатомии этой области мозга [5, 8, 9, 11, 14]. Помимо этого, в настоящее время показано, что островок играет ключевую роль во многих процессах — от висцеросенсорных и восприятия боли до мотивационных, когнитивного контроля речи и эмоций [15—18]. T. Wager назвал островок ключом, соединяющим мышление и аффективную сферу, а A. Craig считал, что передняя часть островка, получающая богатую интерорецепцию и имеющая мощные связи с лимбическими структурами, ответственна за самосознание [19].
В нашей работе мы делали акцент на морфологических особенностях извилин островка и его покрышек, специфике сосудистой системы островковой области с позиций двух основных доступов, используемых для подхода к островку: транссильвиевого и транскортикального.
В классических работах островок описывается как пятая доля головного мозга, по форме напоминающая пирамиду и отграниченная от окружающих ее лобной, теменной и височной долей периинсулярными бороздами. Большинство авторов [5, 11, 14] выделяют переднюю, верхнюю и заднюю периинсулярные борозды. Несколько иной взгляд изложен в работе A. Afif и соавт. [13], где островок представляется в виде трапеции, а авторы описывают 4 периинсулярные борозды: переднюю, верхнюю, заднюю и нижнюю. Исследуя наш анатомический материал, мы придерживались описания передней, верхней и задней периинсулярных борозд.
Как известно, островковая доля кровоснабжается из многочисленных перфорирующих артерий, отходящих от лежащих на ней сосудов М2 сегмента средней мозговой артерии. Однако возникает важный практический вопрос: можно ли их коагулировать во время удаления опухоли? Насколько глубоко данные артерии распространяются в медиальном направлении и где заканчивается зона их кровоснабжения?
Ряд авторов приводят данные о наличии длинных, отличающихся по диаметру перфорирующих артерий, отходящих от М2 сегмента и встречающихся преимущественно в задней части островковой доли.
Эти артерии до нашего исследования описаны только в трех работах, а впервые их описал G. Varnavas и соавт. [14], который обнаружил перфорирующие артерии большего диаметра в верхних отделах задней доли островка в четверти исследуемых гемисфер. Зона кровоснабжения этих артерий не уточнялась.
N. Tanriover и соавт. [11] описали перфорирующие артерии большего диаметра не только в верхне-задней части островка, но и в нижних отделах задней доли.
U. Ture и соавт. [8] указывают, что приблизительно 85—90% инсулярных (отходящих от М2 сегмента) артерий — короткие и кровоснабжают только кору островковой доли и крайнюю капсулу, 10% артерий имеют среднюю длину и доходят до ограды и наружной капсулы, а 3—5% артерий — длинные (встречаются в задней доле островка), кровоснабжающие лучистый венец. Повреждение последних во время резекции опухолей островковой доли может привести к гемипарезу.
Исследуя наш материал, мы обнаружили перфоранты М2 сегмента большего диаметра только в верхних отделах задних длинных извилин, при этом лишь в 2 (11%) гемисферах они кровоснабжали лучистый венец. Во всех остальных случаях они разветвлялись не далее латеральной части скорлупы. Следовательно, медиальной границей зоны кровоснабжения инсулярных артерий является наружная капсула, за исключением верхнезадних отделов островка, где в небольшом количестве случаев перфорирующие артерии доходят до лучистого венца.
Поскольку глиальные опухоли островковой доли кровоснабжаются из перфорантов М2 сегмента, одним из этапов удаления опухоли [1] является ее деваскуляризация путем коагуляции перфорантов М2. Однако, учитывая результаты анатомического исследования, предполагаем, что в задних отделах островка эта стадия доступа (если будет коагулирован большой перфорирующий сосуд) может приводить к ишемическому повреждению лучистого венца и, как следствие, к неврологическому дефициту.
Сохранение лентикулостриарных артерий является одной из сложнейших задач хирургии островковой доли мозга, а повреждение данных артерий рассматривается как главная причина стойкого неврологического дефицита [1, 20]. В связи с этим самая латеральная лентикулостриарная артерия приобретает важное значение как интраоперационный ориентир [21], доступный только при транссильвиевом доступе и позволяющий определить латеральную границу переднего продырявленного вещества. Не менее важным ориентиром во время удаления глиальных опухолей из островка является и его порог, также хорошо распознаваемый при транссильвиевом доступе. По результатам нашего анатомического исследования, место входа самой латеральной лентикулостриарной артерии в переднее продырявленное вещество расположено на расстоянии 16 мм от средины порога островка (что примерно соответствует результатам, полученным N. Tanriover и соавт. — 15,3 мм [11]), а средняя длина участка латеральных лентикулостриарных артерий от места отхождения от М1 сегмента до входа в переднее продырявленное вещество равняется 4 мм.
Отличительной особенностью островка является отсутствие выхода коры доли на поверхность мозга, что затрудняет прямой хирургический доступ к ней. Островок скрыт расположенными над и под ним частями височной, лобной и теменной долей — покрышками. В литературе встречаются различия в обозначении покрышек островка. Ряд авторов выделяют три покрышки: лобную, теменную и височную [11] (или лобноорбитальную, лобно-теменную и височную [5]), другие описывают всего две — лобно-теменную и височную [12]. На наш взгляд, оптимальным является выделение лобной, теменной и височной покрышек, так как в этом случае название и границы покрышек совпадают с долями, в которых они расположены.
Однако варианты обозначения покрышек не имеют принципиального (практического) значения, в отличие от особенностей их строения в передних/задних и верхних/нижних отделах, что определяет различную доступность участков островковой доли при транскортикальном и транссильвиевом доступе.
Островковую долю, на наш взгляд, естественным образом можно разделить на несколько отделов (см. рис. 13). Центральная борозда делит островок на переднюю и заднюю доли, в каждой из которых верхняя часть располагается под лобной/теменной покрышкой, а нижняя часть — под височной. Таким образом, островок разбивается на 4 отдела: передневерхний
,
передненижний
,
задневерхний
,
задненижний.
Мы также считаем целесообразным, в связи с анатомической близостью к переднему продырявленному веществу, выделение в передненижнем отделе
порога
островка.
Толщина покрышек над передней долей островка меньше, чем над задней, а высота лобной и теменной покрышек больше, чем высота височной. Поэтому глубина операционной раны в передних отделах островка меньше, чем в задних.
Помимо этого, ось planum polare, который покрывает передненижний отдел островка, в отличие от всех остальных покрышек, ориентирована под острым углом к сагиттальной плоскости и отклонена латерально (см. рис. 4), что вместе с ретракцией кверху треугольной части увеличивает свободное пространство сильвиевой щели на этом уровне и облегчает ретракцию при транссильвиевом доступе к передненижним отделам островка.
Поэтому, моделируя на анатомических препаратах транссильвиевый доступ с учетом морфологии покрывающих островок покрышек мозга, мы пришли к выводу, что нижние отделы доли более доступны, чем верхние (по причине крайне неудобного верхнезаднего угла атаки и большей высоты лобной и теменной покрышек в сравнении с височной).
Доступность передневерхнего и задневерхнего отделов также отличается. Несмотря на то что глубина раны при доступе к передневерхним отделам островка меньше, чем к задневерхним (см. рис. 3 и 7), расстояние до верхней периинсулярной борозды (длина передневерхнего и задневерхнего отделов, расположенных под лобной и теменной покрышками (см. рис. 3, расстояние В) больше в передневерхнем отделе доли, что приводит к тому, что при транссильвиевом доступе передневерхние и задневерхние отделы становятся одинаково малодоступными. Меньшую толщину покрышек в передневерхнем отделе нивелирует большее расстояние до верхней периинсулярной борозды (см. рис. 3, расстояние В), что делает данный отдел наименее доступным при транссильвиевом доступе.
Таким образом, наиболее доступными при транссильвиевом доступе являются нижние зоны островка (включая порог), а наименее доступными — верхние отделы. Поэтому при локализации опухоли в данных отделах островковой доли может быть рекомендован транскортикальный доступ, который, в отличие от транссильвиевого, не требует значительной ретракции мозгового вещества и обеспечивает больший хирургический коридор.
При моделировании транскортикального доступа единственным отличием в доступности отделов островка была большая глубина операционной раны в задних отделах в сравнении с передними. Так как при транскортикальном доступе производится резекция части покрышки, проекционно расположенной над пораженным опухолью отделом островка, угол атаки (а следовательно, и доступность) на верхние и нижние отделы доли, в отличие от транссильвиевого доступа, не отличается.
Транскортикальный доступ, вне зависимости от отдела островковой доли, обеспечивает больший хирургический обзор и рабочее пространство в сравнении с транссильвиевым, однако при расположении опухоли в пороге островка он не предоставляет надежного проксимального контроля лентикулостриарных артерий, поэтому при локализации опухоли в данной зоне может быть рекомендован транссильвиевый доступ.