Центростремительные волокна
Афферентные нервные пути классифицируются на пути бессознательной и сознательной чувствительности. Именно при помощи них обеспечивается связь между всеми интеграционными центрами, расположенными в головном мозге. К примеру, они обеспечивают прямую связь между мозжечком и корой полушарий большого мозга.
Основные афферентные пути ЦНС сознательной общей чувствительности – это волокна болевой, температурной и тактильной чувствительности, а также сознательной проприоцептивной. Основные бессознательные проводящие пути общей чувствительности – это передний и задний спинно-мозжечковые. К специальным проводящим относятся вестибулярный, слуховой, вкусовой, обонятельный и зрительный.
ДВИГАТЕЛЬНЫЕ (эфферентные) ПРОВОДЯЩИЕ ПУТИ
Эфферентные пути проводят нервный импульс из различных центров головного мозга к рабочему органу. У человека прямые эфферентные пути начинаются лишь из коры больших полушарий, где располагаются тела так называемых центральных нейронов.
Эти пути являются проводниками
сознательных двигательных импульсов
к скелетной мускулатуре и объединяются понятием
пирамидная система.
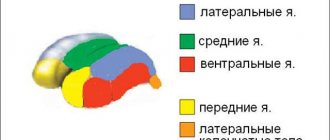
Промежуточные ядра ствола (красное ядро, ядра крыши среднего мозга, ядра ретикулярной формации, ядро оливы, латеральное преддверное ядро) рассматриваются как начало эфферентных проводящих путей, которые посылают бессознательные двигательные импульсы на скелетную мускулатуру и входят в состав
экстрапирамидной системы. Мозжечок осуществляет связь со спинным мозгом через указанные стволовые ядра. Все нисходящие пути заканчиваются на телах и дендритах нейронов двигательных ядер передних рогов серого вещества спинного мозга или двигательных ядер черепных нервов (периферические нейроны).
Таким образом, все эфферентные проводящие пути подразделяются на две группы:
1. Пирамидные пути.
2. Экстрапирамидные пути.
ПИРАМИДНЫЕ ПУТИ
Пирамидные пути проводят сознательные (волевые) двигательные импульсы, а также тормозные импульсы от коры полушарий большого мозга к нейронам двигательных ядер черепных нервов и к нейронам двигательных ядер передних рогов серого вещества спинного мозга.
Для пирамидных путей характерно:
1. Наличие 2-х нейронов.
2. I-й нейрон – пирамидные клетки Беца (V слой коры предцентральной извилины).
3. В полушарии головного мозга волокна проходят в составе лучистого венца и внутренней капсулы, занимая колено и передние 2/3 задней её ножки.
4. В мозговом стволе волокна следуют в его вентральной части, переходя последовательно из ножек мозга в мост и продолговатый мозг.
5. На границе со спинным мозгом 80% волокон, переходя на другую сторону, образуют нижний двигательный перекрест (decussatio pyramidum).
6. В спинном мозге пирамидные пути занимают его передние и боковые канатики.
7. Тела II-х нейронов располагаются в двигательных ядрах передних рогов спинного мозга или в двигательных ядрах черепных нервов.
В зависимости от места назначения пирамидные пути подразделяются на две группы:
• корково-ядерный путь, tractus corticonuclearis (corticobulbaris);
• корково-спинномозговой путь, tractus corticospinalis (pyramidalis) .
Волокна тактильной, температурной и болевой чувствительности
Данный путь берет свое начало с рецепторов в эпителии, импульсы от которых поступают в клетки спинномозгового узла, а после этого в спинной мозг, к ядрам зрительного бугра. Затем к коре постцентральной извилины, в которой и происходит их полный анализ. В данный проводящий путь вовлечено три тракта:
- Таламо-корковый.
- Ганглиоспинномозговой.
- Боковой спинно-таламический тракт, который проходит в боковом канатике спинного мозга и покрышке ствола головного мозга.
За получение тактильных ощущений в лицевой части головы и изменений температуры тела отвечает тройничный нерв. При его повреждении у человека начинаются сильные боли в области лица, которые то исчезают, то вновь появляются. Тройничный нерв проходит через шейный отдел, там, где перекрещиваются моторные волокна кортикоспинального тракта. Аксоны чувствительных нейронов тройничного нерва проходят через одну из частей продолговатого мозга. Через эти аксоны в мозг поступает информация о болевых ощущениях в ротовой полости, зубах, а также в верхней и нижней челюстях.
Закономерности строения афферентных проекционных проводящих путей.
1) Начало каждого пути предст-но рецепторами, заложенными в коже, подкожной ткани или глубоких частях тела.
2) Первый нейрон у всех афферентных проводящих путей нах-ся вне ЦНС, в спинномозговых ганглиях.
3) Второй нейрон локализуется в ядрах спинного или продолговатого мозга.
4) Все восходящие проводящие пути проходят в дорсальной части мозгового ствола.
5) Третий нейрон у путей, направляющихся в кору полушарий большого мозга, расп-ся в ядрах таламуса, а у мозжечковых путей в коре мозжечка.
6) Пути, приносящие импульсы в кору большого мозга, имеют один перекрест, совершаемый отростками 2-го нейрона; благодаря этому каждая половина тела проецируется на противоположное полушарие большого мозга.
7) Мозжечковые пути или не имеют ни одного перекреста, или перекрещиваются дважды, так что каждая половина тела проецируется на кору одноименной половины мозжечка.
 Пути, соединяющие мозжечок с корой полушарий большого мозга, являются перекрещенными.
Пути, соединяющие мозжечок с корой полушарий большого мозга, являются перекрещенными.
9) Корковый центр всех обще чувствительных проводящих путей локализуется в 4 слое коры постцентральной извилины.
Нисходящие (эфферентные) проекционные проводящие пути проводят импульсы от коры, подкорковых центров к нижележащим отделам, к ядрам мозгового ствола и к двигательным ядрам спинного мозга. Эти пути подразделяются: пирамидные, экстрапирамидные
.
Пирамидный путь
связывает нейроны 5 слоя двигательной области коры непосредственно с двигательными ядрами спинного мозга и черепных нервов Он передает мышцам сигналы произвольных движений, регулируемых корой большого мозга. Пирамидные пути служат для осознанного (волевого) управления скелетной мускулатурой.
Экстрапирамидные пути
предст-ны многозвенными нисходящими путями, несут импульсы от коры через подкорковые центры к двигательным ядрам черепных нервов или передних рогов спинного мозга, а затем к скелетным мышцам.
Посредством экстрапирамидной системы осущ-ся регуляция непроизвольных движений, автомат-их двигательных актов, мышечного тонуса, а также движений, выражающих эмоции
Закономерности строения эфферентных проводящих путей
1) Первый нейрон всех эфферентных проводящих путей локализуется в 5 слое предцентральной извилины и парацентральной дольки коры большого мозга.
2) Эфферентные проекционные пути занимают переднюю ножку, колено и переднюю часть задней ножки внутренней капсулы, проходят в основании ножек мозга и моста.
3) Все эфферентные пути заканчиваются в ядрах двигательных черепных нервов и в передних рогах спинного мозга, где расп-ся последний, двигательный нейрон.
4) Эфферентные пути образуют полный или частичный перекрест, вследствие чего импульсы из коры полушария большого мозга передаются мышцам противоположной половины тела.
Проекционные чувствительные проводящие пути
Волокна сознательной общей чувствительности
Этот путь проводит через себя все виды общей чувствительности от головы до шеи. Рецепторы начинают свой путь в мышцах и коже, проводят импульсы к чувствительным ганглиям и переходят в ядра тройничного нерва. Далее путь переходит на зрительные бугры, а потом распространяется на клетки постцентральной извилины. При этом включается три основных тракта:
- таламокорковый;
- ганглиоядерный;
- ядерно-таламический.
Волокна сознательной проприоцептивной чувствительности
Этот путь берет свое начало своими рецепторами в сухожилиях, надкостнице, мышцах и связках, а также в суставных сумках. При этом проводится полная информация о вибрациях, положении тела, степени расслабления и сокращения мышц, давлении и весе. Нейроны данного пути располагаются в спинномозговых узлах, ядрах клиновидного и тонкого бугорков продолговатого мозга, зрительного бугра промежуточного мозга, в котором затем начинается переключение импульсов. Информация анализируется и заканчивает свой путь в центральной извилине коры большого полушария мозга. Этот путь включает в себя три тракта:
- Таламокорковый, который заканчивается в проекционном центре, то есть в центральной извилине головного мозга.
- Тонкий и клиновидный пучки, проходящие в заднем канатике спинного мозга.
- Бульбарно-таламический тракт, проходящий в покрышке ствола головного мозга.
Восходящие (афферентные) пути спинного и головного мозга
Восходящие (афферентные) пути, начинающиеся в спинном мозге
Тела первых нейронов – проводников всех видов чувствительности к спинному мозгу – лежат в спинномозговых узлах. Аксоны клеток спинномозговых узлов в составе задних корешков вступают в спинной мозг и делятся на две группы: медиальную, состоящую из толстых, более миелинизированных волокон, и латеральную, образуемую тонкими, менее миелинизированными волокнами.
Медиальная группа волокон заднего корешка направляется в задний канатик белого вещества, где каждое волокно делится Т-образно на восходящую и нисходящую ветви. Восходящие ветви, следуя кверху, вступают в контакт с клетками серого вещества спинного мозга в студенистом веществе и в заднем роге, а часть их доходит до продолговатого мозга, образуя тонкий и клиновидный пучки, fasciculi gracilis et cuneatus (см. рис. 880, 881, 882), спинного мозга.
Нисходящие ветви волокон направляются вниз и вступают в контакт с клетками серого вещества задних столбов на протяжении шести – семи нижележащих сегментов. Часть этих волокон образует в грудном и шейном отделах спинного мозга пучок, имеющий на поперечном сечении спинного мозга вид запятой и располагающийся между клиновидным и тонким пучками; в поясничном отделе – вид медиального тяжа; в крестцовом отделе – вид овального пучка заднего канатика, примыкающего к медиальной поверхности тонкого пучка.
Латеральная группа волокон заднего корешка направляется в краевую зону, а затем в задний столб серого вещества, где вступает в контакт с расположенными в нем клетками заднего рога.
Волокна, отходящие от клеток ядер спинного мозга, направляются кверху частично по боковому канатику своей стороны, а частично переходят в составе белой спайки на противоположную сторону спинного мозга и также направляются кверху в боковом канатике.
К восходящим путям (см. рис. 880, 881, 882), начинающимся в спинном мозге, относят следующие:
- Задний спинно-мозжечковый путь, tractus spinocerebellaris dorsalis [posterior], – прямой мозжечковый путь, проводит импульсы от рецепторов мышц и сухожилий к мозжечку. Тела первых нейронов лежат в спинномозговом узле, тела вторых нейронов – на всем протяжении спинного мозга в грудном столбе (грудном ядре) заднего рога. Длинные отростки вторых нейронов идут кнаружи; достигнув задненаружного отдела спинного мозга той же стороны, заворачивают вверх и поднимаются вдоль бокового канатика спинного мозга, а затем по нижней мозжечковой ножке следуют к коре червя мозжечка.
- Передний спинно-мозжечковый путь, tractus spinocerebellaris ventralis [anterior], проводит импульсы от рецепторов мышц и сухожилий к мозжечку. Тела первых нейронов лежат в спинномозговом узле, а вторых нейронов – в медиальном ядре промежуточной зоны и посылают часть своих волокон через белую спайку в боковые канатики противоположной стороны, а часть – в боковые канатики своей стороны. Указанные волокна достигают передненаружных отделов боковых канатиков, располагаясь кпереди от заднего спинно-мозжечкового пути. Здесь волокна заворачивают вверх, идут по спинному, а затем по продолговатому мозгу и, пройдя мост, по верхним мозжечковым ножкам, совершив второй перекрест, достигают червя мозжечка.
- Спиннооливный путь, tractus spinoolivaris, берет начало от клеток задних рогов серого вещества. Аксоны этих клеток перекрещиваются и поднимаются вблизи поверхности спинного мозга на границе бокового и переднего канатиков, заканчиваясь в ядрах оливы. Волокна этого пути несут информацию от кожных, мышечных и сухожильных рецепторов.
Рис. 952. Проводящие пути внутренней капсулы и ножек мозга (полусхематично).Рис. 951. Капсулы и ход проводящих путей через внутреннюю капсулу (полусхематично).
- Передний и латеральный спинно-таламические пути, tractus spinothalamici ventralis [anterior] et lateralis (см. рис. 947), проводят импульсы болевой, температурной (латеральный путь) и тактильной (передний путь) чувствительности. Тела первых нейронов лежат в спинномозговых ганглиях. Отростки вторых нейронов от клеток собственного ядра заднего рога направляются через белую спайку в передний и боковой канатики противоположной стороны. Поднимаясь вверх, волокна этих путей проходят в задних отделах продолговатого мозга, моста и ножек мозга и достигают таламуса в составе спинномозговой петли, lemniscus spinalis. В таламусе залегают тела третьих нейронов этих путей, а их отростки направляются к коре головного мозга в составе центральных таламических лучистостей через заднюю ножку внутренней капсулы (рис. 951, 952).
- Спинно-ретикулярный путь, tractus spinoreticularis, составляют волокна, которые проходят в составе спинно-таламических путей, не перекрещиваются и образуют двусторонние проекции на все отделы стволовой ретикулярной формации.
- Спинно-покрышечный путь, tractus spinotectalis, вместе со спинно-таламическим путем проходит в боковых канатиках спинного мозга и заканчивается в пластинке крыши среднего мозга.
- Тонкий пучок, fasciculus gracilis, и клиновидный пучок, fasciculus cuneatus (см. рис. 947), проводят импульсы от мышц, суставов и рецепторов тактильной чувствительности. Тела первых нейронов этих путей локализуются в соответствующих спинномозговых узлах. Аксоны идут в составе задних корешков и, вступив в задние столбы спинного мозга, принимают восходящее направление, достигая ядер продолговатого мозга.
Тонкий пучок занимает медиальное положение и проводит соответствующие импульсы от нижних конечностей и нижних частей туловища – ниже 4-го грудного сегмента.
Клиновидный пучок образуется волокнами, начинающимися от клеток всех спинномозговых узлов, лежащих выше 4-го грудного сегмента.
Достигнув продолговатого мозга, волокна тонкого пучка вступают в контакт с клетками ядра этого пучка, лежащего в бугорке тонкого ядра; волокна клиновидного пучка заканчиваются в клиновидном бугорке. Клетки обоих бугорков являются телами вторых нейронов описываемых путей. Их аксоны – внутренние дугообразные волокна, fibrae arcuatae internae, – направляются вперед и вверх, переходят на противоположную сторону и, образуя перекрест медиальных петель (чувствительный перекрест), decussatio lemniscorum medialium (decussatio sensoria), с волокнами противоположной стороны, идут в составе медиальной петли, lemniscus medialis.
Достигнув таламуса, эти волокна вступают в контакт с его клетками – телами третьих нейронов пути, которые посылают через внутреннюю капсулу свои отростки к коре головного мозга.
Спиномозговые волокна
Афферентные пути спинного мозга образуются при помощи аксонов, или, как их еще называют по-другому, окончания нейронов. Аксоны находятся только в спинном мозге и не выходят за его пределы, а также создают связь между всеми сегментами органа. Атомическое строение данных волокон состоит в том, что длина аксонов довольно большая и соединяется с другими нервными окончаниями. От рецепторов к центральной нервной системе за счет афферентных проводящих путей спинного и головного мозга проводятся нервные сигналы. Все нервные волокна, расположенные по всей длине спинного мозга, участвуют в данном процессе. Сигнал к органам проводится от разных отделов ЦНС и между нейронами. Беспрепятственное прохождение сигнала от периферии к центральной нервной системе достигается при помощи проводящих путей спинного мозга.
Как они работают вместе и чем отличаются
Афферентные нейроны обычно имеют два аксона, которые передают электрохимические сигналы в позвоночный столб или мозг. Оказавшись там, сигнал проходит через сеть интернейронов и через эфферентный нейрон. Афферентно-эфферентные пары нейронов, которые проходят через позвоночник, управляют рефлексами (такими, как реакция коленного рефлекса).
Готовые работы на аналогичную тему
Курсовая работа Афферентные и эфферентные нервные проводники и их роль в психологии 410 ₽ Реферат Афферентные и эфферентные нервные проводники и их роль в психологии 240 ₽ Контрольная работа Афферентные и эфферентные нервные проводники и их роль в психологии 240 ₽
Получить выполненную работу или консультацию специалиста по вашему учебному проекту Узнать стоимость
Афферентные нейроны предназначены для реагирования на различные раздражители. Например, афферентный нейрон, предназначенный для реакции на тепло, обнаруживает избыточное тепло и посылает импульс через центральную нервную систему. Затем эфферентный нейрон заставляет мышцы сокращаться, чтобы отвести тело от жары. Кожа имеет сенсорные рецепторы для тепла, холода, удовольствия, боли и давления.
Афферентные нейроны имеют круглые и гладкие клеточные тела, в то время как эфферентные нейроны имеют спутниковые тела. Афферентные нейроны обнаруживаются в периферической нервной системе, а эфферентные нейроны располагаются в центральной нервной системе. Аксоны в афферентных нейронах движутся от ганглиев (скопление нервных клеток, в которых находятся афферентные и эфферентные нейроны) к спинному мозгу. Длинный аксон фактически связан с эфферентным нейроном.
Афферентные нейроны имеют один длинный миелинизированный дендрит, тогда как эфферентные нейроны имеют более короткие дендриты. Дендрит в афферентном нейроне — это то, что отвечает за передачу нервных импульсов от рецепторов к телу клетки, в то время как в эфферентном нейроне импульсы проходят через дендрит и выходят через нервно-мышечное соединение, которое образуется между эффекторами и аксоном.
Задний и передний спиномозжечковые тракты
Афферентные пути мозжечка относятся к бессознательным и берут свое начало в боковом канатике спинного мозга, а оттуда уже проводят информацию о том, в каком состоянии находятся органы опорно-двигательного аппарата. Передний спиномозжечковый тракт поступает в мозжечок через верхнюю ножку, в связи с этим он проходит через покрышку продолговатого мозга, среднего мозга и моста. Задний спиномозжечковый тракт проходит в продолговатом мозге и проникает через нижнюю ножку.
Эти два тракта передают в мозжечок информацию от связок, суставных сумок, рецепторов мышц, сухожилий, надкостницу. Именно они отвечают за поддержание равновесия и координацию движений человека, поэтому роль их в организме очень важна.
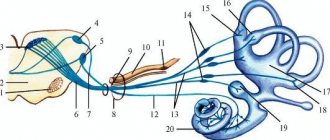
Слуховые волокна
Данный путь проводит информацию от рецепторов Кортиева органа, который располагается во внутреннем ухе. Нервные импульсы поступают в мост, содержащий в себе слуховые ядра по волокнам преддверно-улиткового нерва. Через слуховые ядра информация передается на ядра трапециевидного тела. После этого импульсы поступают на подкорковые центры слуха, которые включают в себя зрительный бугор, нижние холмики и коленчатые медиальные тела.
В среднем мозге на эти слуховые раздражители возникают возвратные реакции, при этом афферентные слуховые пути переключаются в ядра таламуса, в которых происходит оценка слуховых раздражителей – они отвечают за движения, происходящие непроизвольно: ходьба, бег. От коленчатых тел начинает исходить слуховая лучистость – этот тракт проводит импульсы от внутренней капсулы до проекционного центра слуха. Только здесь уже начинает происходить оценка звуков. В задней части височной извилины располагается ассоциативный слуховой центр. Именно в нем все звуки начинают восприниматься как слова.
Вкусовые анализаторы
Импульсы афферентного пути вкусовых анализаторов развиваются от рецепторов корня языка, входящих в состав языкоглоточных нервов и располагающихся на языке, которые входят в состав лицевого нерва. Импульсы от них поступают в продолговатый мозг, а потом к ядрам лицевого и языкоглоточного нерва. Наименьшая часть всей получаемой информации от этих импульсов доставляется в мозжечок, образуя тем самым ядерно-мозжечковый путь, и обеспечивает рефлекторную регуляцию тонуса мышц языка, головы и глотки. Большая часть информации поступает в зрительные бугры, после чего импульсы достигают крючка височной доли, в котором происходит их сознательный анализ.
Ассоциативные пути, короткие и длинные.
Ассоциативные пути являются связующим звеном между последним нейроном афферентного пути и первым нейроном эфферентного пути.
Ассоциативные пути представляют собой цепи вставочных нейронов, соединяющие различные участки коры в пределах одного полушария. В СМ ассоциативные пути связывают соседние сегменты.
Выделяют короткие и длинные ассоциативные пути.
Короткие ассоциативные волокна соединяют участки коры соседних извилин, не выходя за пределы доли полушария мозга. Это дугообразные волокна, располагающиеся поверхностно под корой — на дне борозд. Сюда же относят волокна, связывающие клетки соседних ядер ствола мозга.
Длинные ассоциативные волокна располагаются под слоем коротких ассоциативных волокон в полушариях большого мозга. Они связывают участки коры различных долей одного полушария.
К длинным ассоциативным волокнам относят: 1. Пояс. Это группа нервных волокон, располагающаяся в глубине сводчатой извилины. Соединяет участки коры лобной, затылочной и височной долей в области медиальной поверхности полушария большого мозга. В онтогенезе пояс развивается раньше других ассоциативных путей. 2. Верхний продольный пучок. Локализуется под верхне-латеральной поверхностью полушария большого мозга, латеральнее лучистого венца.
Его волокна соединяют участки коры нижних отделов лобной доли, нижней теменной дольки, височной и затылочной долей. Формирование в онтогенезе верхнего продольного пучка связано с развитием корковых концов кожного, двигательного, слухового и зрительного анализаторов.
3. Нижний продольный пучок. Располагается под нижне-медиальной поверхностью полушария большого мозга, вдоль наружной стенки заднего и нижнего рогов бокового желудочка.
Пучки волокон соединяют участки коры затылочной и височной долей. Нижний продольный пучок развивается раньше, чем верхний и обеспечивает связь коркового конца зрительного анализатора с вегетативными центрами, а значит — их сочетанные действия. 4. Крючковидный пучок. Располагается на нижне-латеральной поверхности полушария большого мозга. Соединяет участки коры лобной, височной и затылочной долей полушария. К длинным ассоциативным путям относят волокна, проходящие в составе терминальной полоски, мозговой полоски таламуса, дорсального и медиального продольных пучков.
В СМ ассоциативные волокна располагаются вокруг СВ в виде узкой полосы.
Это собственные пучки, которые относят к коротким ассоциативным путям. Они входят в состав местных рефлекторных дуг, связывая между собой сегменты СМ, а также чувствительные нейроны спинномозговых узлов с двигательными нейронами передних рогов СМ.
В онтогенезе ассоциативные пути формируются значительно позднее по сравнению с комиссуральными и проекционными путями.
Однако, в дальнейшем ассоциативные пути быстро развиваются и преобладают над остальными путями.
Комиссуральные пути. Проекционные пути: а) восходящие (афферентные) системы волокон.
Комиссуральные нервные пути — проводящие пути ЦНС, соединяющие симметричные части полушарий большого мозга или других отделов ЦНС. Комиссуральные (спаечные) пути соединяют между собой участки коры правого и левого полушарий большого мозга и обеспечивают единство деятельности мозга.
Спаечные, то есть комиссуральные пути соединяют как симметричные, так и асимметричные участки коры разных полушарий.
К комиссуральным путям относят: мозолистое тело, переднюю спайку мозга, заднюю спайку мозга, спайку свода.
Мозолистое тело. Среди других комиссур филогенетически наиболее молодое образование.
Состоит из поперечно направленных нервных волокон, соединяющих аналогичные участки новой коры правого и левого полушарий большого мозга друг с другом. Часть волокон, образующих клюв мозолистого тела, связывает между собой оба таламуса и головки хвостатых ядер, обеспечивая синхронную деятельность не только коры правого и левого полушарий, но и подкорки. Передняя спайка мозга. Представлена волокнами, относящимися к проводящим путям обонятельного мозга.
Передняя спайка мозга располагается впереди столбов мозгового свода и состоит из передней и задней частей. Формирование передней части спайки связано с формированием древней коры, а задней части — с формированием новой коры. Высказывается предположение о значении передней спайки для парной деятельности не только обонятельного, но и слухового и зрительного анализаторов. Передняя спайка обеспечивает межполушарную интеграцию импульсов.
Задняя спайка мозга. Располагается в задней стенке III желудочка. Волокна, составляющие спайку, соединяют между собой подушки таламусов. Спайка свода (гиппокампова спайка).Это древнее образование, относящееся к проводящим путям обонятельного мозга. Волокна спайки свода связывают между собой структуры гиппокампа правого и левого полушарий и обеспечивают синхронную работу гиппокампа.
Спайка свода расположена между нижней поверхностью валика мозолистого тела и ножками свода.
Проекционные нервные пути — проводящие пути ЦНС, связывающие кору большого мозга с периферией и проходящие через различные отделы ЦНС. Проекционные нервные пути подразделяются на афферентные (восходящие) и эфферентные (нисходящие).
К восходящим путям бокового столба относятся следующие: спиноталамический, или tractus spino-thalamicus — волокна вторых нейронов болевого и температурного, отчасти тактильного чувства, перешедшие в противоположный боковой столб после перекреста в передней серой спайке.
Помимо простейших рефлекторных дуг, которые возникают в пределах одного сегмента СМ, также существуют межсегментные рефлекторные восходящие и нисходящие пути.
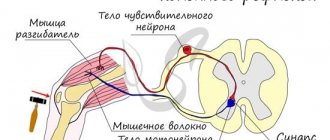
Рассмотрим восходящие пути. При выполнении коленного рефлекса мышца-разгибатель сокращается, и нога непроизвольно разгибается. Сигналы от чувствительного нейрона, сигнализирующие о том, что нога изменила положение, идут вверх по коллатерали чувствительного нейрона.
Эта коллатераль проходит в БВ СМ к таламусу. Здесь происходит фильтрация приходящих сигналов. Например, если сигнал одиночный и слабый, то он просто не проходит.
Такая система позволяет регулировать прохождение сигналов к коре и дает возможность реагировать на более важные сигналы с наибольшей быстротой. После прохождения таламуса нервный импульс приходит к нейронам соматосенсорной коры больших полушарий конечного мозга. При этом у человека возникает ощущение, что нога разогнулась.
Для того, чтобы вернуть ее в исходное положение, нервный импульс передается в моторную кору больших полушарий, где происходит построение программ движений. Межсегментные восходящие пути позволяют нам контролировать произвольные движения, запускаемые на разных уровнях СМ.
Зрительные анализаторы
Афферентные проводящие пути ЦНС зрительного анализатора начинаются от колбочек и палочек сетчатки глазного яблока. Импульсы поступают в зрительный перекресток в составе зрительных нервов, а затем по тракту направляются к подкорковым центрам головного мозга, то есть в зрительный бугор, коленчатые латеральные тела и верхние холмики, расположенные в средней части мозга.
В среднем мозге на эти раздражители возникает ответная реакция, при этом в ядрах таламуса начинается бессознательная оценка импульсов, которые обеспечивают непроизвольные движения, воспроизводимыми человеком. Основными такими бессознательными движениями являются бег и ходьба. В проекционном центре зрения или в шпорную борозду затылочной доли головного мозга импульсы поступают по зрительной лучистости от коленчатых тел, находящихся в составе внутренней капсулы, после чего начинается полный анализ поступающих данных. В коре, которая прилегает к шпорной борозде, находит свое место расположения центральная часть, отвечающая за зрительную память, которую еще называют ассоциативным зрительным центром.
Восходящие (афферентные) пути, начинающиеся в стволе головного мозга
В стволе головного мозга начинаются медиальная петля, тройничная петля, восходящий путь слухового анализатора, зрительная лучистость, таламические лучистости.
1. Медиальная петля как продолжение тонкого и клиновидного пучков описана ранее.
2. Тройничная петля, lemniscus trigeminalis, образована отростками нервных клеток, составляющих чувствительные ядра тройничного нерва (V пара), лицевого нерва (VII пара), языкоглоточного нерва (IX пара) и блуждающего нерва (X пара).
К чувствительным ядрам тройничного нерва подходят аксоны афферентных нейронов, залегающих в тройничном узле. К общему чувствительному ядру трех других нервов – ядру одиночного пути – подходят аксоны афферентных нейронов, залегающих в узле коленца (VII пара) и в верхних и нижних узлах IX и X пар нервов. В перечисленных узлах локализуются тела первых нейронов, а в чувствительных ядрах – тела вторых нейронов пути, по которому передаются импульсы от рецепторов головы.
Волокна тройничной петли переходят на противоположную сторону (часть волокон следует на своей стороне) и достигают таламуса, где заканчиваются в его ядрах.
Нервные клетки таламуса являются телами третьих нейронов восходящих путей черепных нервов, аксоны которых в составе центральных таламических лучистостей через внутреннюю капсулу направляются к коре головного мозга (постцентральная извилина).
3. Восходящий путь слухового анализатора имеет в качестве первых нейронов клетки, залегающие в узле улитковой части преддверно-улиткового нерва. Аксоны этих клеток подходят к клеткам переднего и заднего улитковых ядер (вторые нейроны). Отростки вторых нейронов, переходя на противоположную сторону, образуют трапециевидное тело, а затем принимают восходящее направление и получают название латеральной петли, lemniscus lateralis. Эти волокна заканчиваются на телах третьих нейронов слухового пути, залегающих в латеральном коленчатом теле. Отростки третьих нейронов образуют слуховую лучистость, radiatio acustica, которая идет от медиального коленчатого тела через заднюю ножку внутренней капсулы к средней части верхней височной извилины.
4. Зрительная лучистость, radiatio optica (см. рис. 951), соединяет подкорковые центры зрения с корой шпорной борозды.
В состав зрительной лучистости входят две системы восходящих волокон:
- коленчато-корковый зрительный тракт, который начинается от клеток латерального коленчатого тела;
- подушково-корковый тракт, начинающийся от клеток ядра, залегающего в подушке таламуса; у человека развит слабо.
Совокупность этих волокон обозначают как задние таламические лучистости, radiationes thalamicae posteriores.
Поднимаясь к коре мозга, обе системы проходят через заднюю ножку внутренней капсулы.
5. Таламические лучистости, radiationes thalamicae (см. рис. 947), образованы отростками клеток таламуса и составляют конечные отделы восходящих путей коркового направления.
В состав таламических лучистостей входят:
- передние таламические лучистости, radiationes thalamicae anteriores, – радиально идущие волокна белого вещества больших полушарий. Они начинаются от верхнего медиального ядра таламуса и направляются через переднюю ножку внутренней капсулы в кору боковой и нижней поверхностей лобной доли. Часть волокон передних таламических лучистостей связывает переднюю группу ядер таламуса с корой медиальной поверхности лобных долей и передней части поясной извилины;
- центральные таламические лучистости, radiationes thalamicae centrales, – радиальные волокна, связывающие вентролатеральную группу ядер таламуса с корой пре- и постцентральной извилины, а также с прилежащими отделами коры лобной и теменной долей. Проходят в составе задней ножки внутренней капсулы;
- нижняя ножка таламуса, pedunculus thalami inferior, содержит радиальные волокна, связывающие подушку таламуса и медиальные коленчатые тела с участками височной хоры;
- задние таламические лучистости (см. ранее).
Обонятельный анализатор
Афферентный путь обонятельного анализатора берет свое начало от рецепторов слизистой оболочки, локализующейся в верхней части носового хода. После этого импульсы направляются к аксонам обонятельных луковиц, и протекают они по волокнам обонятельных нервов. Затем импульсы направляются к проекционному центру обоняния, который находится в районе парагиппокампальной извилины и крючка. Следуют эти импульсы по тракту до коры височной доли головного мозга. К подкорковым центрам, которые располагаются в средней и промежуточной части мозга, направляется большая часть информации, получаемая от обонятельных рецепторов. Подкорковые центры мозга в ответ на обонятельные раздражители обеспечивают рефлекторную регулировку тонуса мышц.
Исходя из этого, можно определить, что основная особенность обонятельных рецепторов заключается в том, что нервные импульсы изначально поступают в кору полушарий головного мозга, а не в подкорковые центры обоняния. В связи с этим, человек сначала чувствует запах, затем начинает оценивать его и только после этого в мозге формируется бессознательная окраска раздражителя на эмоциональном уровне. На весь процесс при этом затрачиваются всего лишь доли секунды.
Интраоперационная идентификация функциональных зон коры и проводящих путей мозга
Еще в 30-е годы прошлого века были предприняты первые шаги в определении локализации корковой функциональной организации в ходе нейрохирургических операций [46, 76]. Изначально использовалась лишь прямая стимуляция коры в ходе удаления опухолей и эпилептогенных зон у больных, находящихся в сознании, что позволяло оценить взаимосвязи сознания, двигательных актов и языковой продукции на этапах хирургического вмешательства. Позднее в 70-х годах XX века в нейрофизиологическую практику вошел мониторинг соматосенсорных вызванных потенциалов (ССВП) [73], и уже совсем недавно, в 90-е годы появились моторные вызванные потенциалы, что позволило более точно локализовать функциональные зоны мозговой коры и проводящие пути.
В 1978 г. был описан феномен реверсии фазы коркового компонента ССВП в области центральных извилин и идентификация центральной борозды [51], который нашел широкое применение в хирургии внутримозговых опухолей [28, 59, 63]. В 1937 г. W. Penfield [76] описал методику прямой стимуляции коры с использованием подачи ритмических (50-60 Гц) биполярных импульсов. Этот подход в основном используется для картирования сенсорных и моторных зон речи у больных в сознании, хотя применяется и у больных, находящихся под общим наркозом [63]. Также в настоящее время применяется методика стимуляции пачками (трейнами) из 4-5 импульсов [90] для картирования и длительного мониторинга корково-подкорковых проводников [17].
Хирургия с пробуждением и прямая электрическая стимуляция коры и проводящих путей являются «золотым стандартом» при резекции глиом, расположенных рядом с речевыми зонами и другими функционально значимыми структурами [36, 38, 42, 74]. В ходе подобных операций функциональное картирование мозга проводится нейрохирургом совместно с нейропсихологом и нейрофизиологом, причем последний осуществляет настройку параметров стимуляции с одновременным контролем состояния спонтанной корковой активности с помощью электрокортикограммы (ЭКоГ) для выявления эпилептической активности. Такой подход позволяет предупреждать развитие судорожного синдрома в ходе продолжительной ритмической электростимуляции. Первым этапом электрическая стимуляция выявляет локализацию корковых зон языковых функций, при этом каждая функционально важная зона маркируется [2, 11, 12].
Зона стимуляции считается функционально значимой, когда наблюдаются речевые нарушения трижды подряд после последующих стимулов, при этом после окончания стимуляции отмечается восстановление речевой (языковой) функции. Тип речевых нарушений верифицируется нейропсихологом, а степень выраженности речевых нарушений оценивается по шкале National Institutes of Health Stroke Scale [92]. Следующим этапом выполняется резекция опухоли с учетом данных картирования, при этом проводится периодическая субкортикальная стимуляция для поиска функционально значимых проводящих путей белого вещества [2].
Таким образом, использование интраоперационного картирования и электрофизиологического мониторинга позволяет хирургам удалить максимальный объем опухоли с минимальным нарушением неврологических функций [31, 54, 61].
4. Нейропсихологические методы интраоперационного исследования
Исследование когнитивных функций в динамике (до и после операции) осуществляется с помощью хорошо подобранного комплекса нейропсихологических тестов, которые должны удовлетворять нескольким критериям: 1) позволять оценить ряд когнитивных функций, при этом тесты должны быть достаточно чувствительные для определения эффекта опухоли и проведенного лечения; 2) процедура тестирования должна быть стандартизирована для обеспечения возможности сравнения результатов динамического обследования пациентов; 3) наличие нормативов проведения тестов позволит объективно оценить выполнение тестов пациентами; 4) тесты должны быть достаточно надежные и малочувствительные к повторному тестированию для получения точных данных при динамическом обследовании; 5) тесты должны иметь альтернативные формы для повторного тестирования; 6) для предотвращения утомления общее время тестирования не должно превышать 30-40 мин.
Комплекс нейропсихологического тестирования подбирается индивидуально для каждого пациента. Поскольку при опухолях часто нарушаются память, внимание, скорость переработки информации, управляющие функции и речь, то соответствующие тесты должны обязательно использоваться при тестировании. В качестве примера можно привести тесты на вербальные ассоциации (ассоциации на заданную букву алфавита и на определенную семантическую категорию), нахождение сходства между двумя понятиями (например, «что общего между яблоком и бананом»), тест следования по маршруту А и Б (позволяет оценить зрительное внимание, распределение и переключение внимания) [65], запоминание списка слов (позволяет оценить слухоречевую память) [55, 60, 83], тест шифровка (позволяет оценить скорость переработки информации) [93], тестом на тонкую моторику [53]. В зависимости от локализации поражения исследование может быть дополнено тестом интеллекта Векслера (WAIS-IV), который позволяет получить суммарную оценку вербального и невербального интеллекта [93]. Тест составной фигуры Рея (Rey Complex Figure Test) позволяет оценить зрительно-конструктивные функции. Используются также тесты на запоминание невербализуемых фигур (Rey Visual Design Learning Test, Rey Complex Figure Test) [65, 89], тесты на речевую и неречевую рабочую память (Digit Span Forward, Digit Span Backward, Spatial Span Forward, Spatial Span Backward) [65, 89], тесты для исследования речи [22].
Для интраоперационного картирования используется небольшой набор относительно несложных тестов, доступных для выполнения в условиях интраоперационного пробуждения. Каждый тест должен включать достаточное количество проб, позволяющих непрерывно тестировать функцию по ходу мозговой стимуляции. В качестве примера приведем несколько проб, используемых для интраоперационной локализации моторных и речевых зон, чаще всего определяемых в ходе подобных операций. Для локализации моторной зоны руки используется проба на сжимание и разжимание кулака, для локализации моторной зоны ноги используется проба на сгибание и разгибание ноги в коленном суставе. Для оценки речевых функций используется проба на называние картинок, при этом называние глаголов является более чувствительным для локализации зоны Брока, чем называние существительных. Порядковый счет в прямом и обратном порядке позволяет оценить плавность речи и переключение.
Таким образом, нейропсихологическое обследование когнитивных функций является важным составляющим комплексного обследования пациентов с опухолями мозга в до- и послеоперационном периоде, определяя как хирургическую тактику, так и последующие реабилитационные мероприятия.
Вестибулярный тракт
Вестибулярный афферентный путь начинается от рецепторов полукружного канала внутреннего уха, маточки и рецепторов, входящих в состав этого органа. Данный тракт в центральной нервной системе отвечает координацию движений и поддержку равновесия при физических и вестибулярных нагрузках.
Афферентные центростремительные проводящие пути и особенность их строения свидетельствуют о том, человеку нужно прилагать массу усилий, чтобы сохранить здоровье и целостность каждого органа по отдельности и вместе взятых. Каждая составляющая данного пути обеспечивает организм всей необходимой информацией, помогает сразу же обрабатывать ее и приводить в исполнение осуществление всех жизненно важных процессов. Это важно в работе всего организма в целом и отдельных органов.