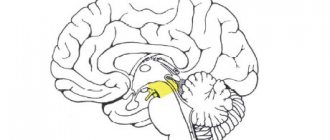
Centripetal fibers
Afferent nerve pathways are classified into unconscious and conscious sensory pathways. It is with their help that communication is ensured between all integration centers located in the brain. For example, they provide a direct connection between the cerebellum and the cerebral cortex.
The main afferent pathways of the central nervous system for conscious general sensitivity are fibers of pain, temperature and tactile sensitivity, as well as conscious proprioceptive. The main unconscious pathways of general sensitivity are the anterior and posterior spinocerebellar. Special conductors include vestibular, auditory, gustatory, olfactory and visual.
MOTOR (efferent) CONDUCTING PATHWAYS
Efferent pathways conduct nerve impulses from various centers of the brain to the working organ. In humans, direct efferent pathways begin only from the cerebral cortex, where the bodies of the so-called central neurons are located.
These pathways are conductors
of conscious motor impulses
to the skeletal muscles and are united by the concept
of the pyramidal system .
The intermediate nuclei of the trunk (red nucleus, nuclei of the roof of the midbrain, nuclei of the reticular formation, olive nucleus, lateral vestibular nucleus) are considered as the beginning of efferent pathways that send unconscious motor impulses to the skeletal muscles and are part of
the extrapyramidal system. The cerebellum communicates with the spinal cord through these stem nuclei. All descending pathways end on the bodies and dendrites of neurons of the motor nuclei of the anterior horns of the gray matter of the spinal cord or the motor nuclei of the cranial nerves (peripheral neurons).
Thus, all efferent pathways are divided into two groups:
1. Pyramid paths.
2. Extrapyramidal tracts.
PYRAMID PATH
The pyramidal tracts conduct conscious (volitional) motor impulses, as well as inhibitory impulses from the cerebral cortex to the neurons of the motor nuclei of the cranial nerves and to the neurons of the motor nuclei of the anterior horns of the gray matter of the spinal cord.
Pyramid paths are characterized by:
1. Presence of 2 neurons.
2. Ist neuron – Betz pyramidal cells (V layer of the cortex of the precentral gyrus).
3. In the cerebral hemisphere, the fibers pass as part of the corona radiata and the internal capsule, occupying the knee and the anterior 2/3 of its posterior legs.
4. In the brain stem, fibers follow in its ventral part, passing successively from the cerebral peduncles to the pons and medulla oblongata.
5. At the border with the spinal cord, 80% of the fibers, passing to the other side, form the lower motor chiasm (decussatio pyramidum).
6. In the spinal cord, the pyramidal tracts occupy its anterior and lateral funiculi.
7. The bodies of II neurons are located in the motor nuclei of the anterior horns of the spinal cord or in the motor nuclei of the cranial nerves.
Depending on the destination, pyramidal paths are divided into two groups:
• corticonuclear pathway, tractus corticonuclearis (corticobulbaris);
• corticospinal tract, tractus corticospinalis (pyramidalis).
Fibers of tactile, temperature and pain sensitivity
This path originates from receptors in the epithelium, impulses from which enter the cells of the spinal ganglion, and then into the spinal cord, to the nuclei of the optic thalamus. Then to the cortex of the postcentral gyrus, where their complete analysis takes place. Three tracts are involved in this pathway:
- Thalamo-cortical.
- Gangliospinal.
- The lateral spinothalamic tract, which runs in the lateral funiculus of the spinal cord and the tegmentum of the brain stem.
The trigeminal nerve is responsible for receiving tactile sensations in the facial part of the head and changes in body temperature. When it is damaged, a person begins to experience severe pain in the facial area, which either disappears or reappears. The trigeminal nerve passes through the cervical spine where the motor fibers of the corticospinal tract cross. The axons of the sensory neurons of the trigeminal nerve pass through one of the parts of the medulla oblongata. Through these axons, the brain receives information about pain in the mouth, teeth, and also in the upper and lower jaws.
Regularities of the structure of afferent projection pathways.
1) The beginning of each path is represented by receptors located in the skin, subcutaneous tissue or deep parts of the body.
2) The first neuron in all afferent pathways is located outside the central nervous system, in the spinal ganglia.
3) The second neuron is localized in the nuclei of the spinal cord or medulla oblongata.
4) All ascending pathways pass in the dorsal part of the brain stem.
5) The third neuron in the pathways heading to the cerebral cortex is located in the nuclei of the thalamus, and in the cerebellar pathways in the cerebellar cortex.
6) The pathways bringing impulses to the cerebral cortex have one crossing, made by the processes of the 2nd neuron; thanks to this, each half of the body is projected onto the opposite hemisphere of the cerebrum.
7) The cerebellar tracts either do not have a single crossover, or cross twice, so that each half of the body is projected onto the cortex of the same half of the cerebellum.
 The pathways connecting the cerebellum to the cerebral cortex are crossed.
The pathways connecting the cerebellum to the cerebral cortex are crossed.
9) The cortical center of all generally sensitive pathways is localized in the 4th layer of the cortex of the postcentral gyrus.
Descending (efferent) projection pathways conduct impulses from the cortex, subcortical centers to the underlying sections, to the nuclei of the brain stem and to the motor nuclei of the spinal cord. These pathways are subdivided: pyramidal, extrapyramidal
.
Pyramid Path
connects neurons of layer 5 of the motor cortex directly with the motor nuclei of the spinal cord and cranial nerves. It transmits signals to the muscles of voluntary movements regulated by the cerebral cortex. The pyramidal tracts serve for conscious (volitional) control of skeletal muscles.
Extrapyramidal tracts
They are presented in multi-link descending pathways, carry impulses from the cortex through the subcortical centers to the motor nuclei of the cranial nerves or anterior horns of the spinal cord, and then to the skeletal muscles.
Through the extrapyramidal system, the regulation of involuntary movements, automatic motor acts, muscle tone, as well as movements expressing emotions is carried out
Regularities of the structure of efferent pathways
1) The first neuron of all efferent pathways is localized in layer 5 of the precentral gyrus and paracentral lobule of the cerebral cortex.
2) Efferent projection pathways occupy the anterior leg, knee and anterior part of the posterior leg of the internal capsule, passing at the base of the cerebral peduncles and pons.
3) All efferent pathways end in the nuclei of the motor cranial nerves and in the anterior horns of the spinal cord, where the last motor neuron is located.
4) Efferent pathways form a complete or partial decussation, as a result of which impulses from the cerebral cortex are transmitted to the muscles of the opposite half of the body.
Projection sensory pathways
Fibers of conscious general sensitivity
This path carries through itself all types of general sensitivity from the head to the neck. The receptors begin their journey in the muscles and skin, conduct impulses to the sensory ganglia and pass into the nuclei of the trigeminal nerve. Next, the path passes to the visual thalamus, and then spreads to the cells of the postcentral gyrus. This includes three main paths:
- thalamocortical;
- ganglionuclear;
- nuclear-thalamic.
Fibers of conscious proprioceptive sensitivity
This pathway originates with its receptors in the tendons, periosteum, muscles and ligaments, as well as in the joint capsules. In this case, complete information is provided about vibrations, body position, degree of relaxation and muscle contraction, pressure and weight. The neurons of this pathway are located in the spinal ganglia, the nuclei of the sphenoid and thin tubercles of the medulla oblongata, and the optic thalamus of the diencephalon, in which the switching of impulses then begins. Information is analyzed and ends its path in the central gyrus of the cerebral cortex. This path includes three tracts:
- Thalamocortical, which ends in the projection center, that is, in the central gyrus of the brain.
- Thin and wedge-shaped bundles passing in the posterior cord of the spinal cord.
- The bulbar-thalamic tract passes through the tegmentum of the brain stem.
Ascending (afferent) pathways of the spinal cord and brain
Ascending (afferent) pathways starting in the spinal cord
The bodies of the first neurons - conductors of all types of sensitivity to the spinal cord - lie in the spinal ganglia. The axons of the cells of the spinal ganglia as part of the dorsal roots enter the spinal cord and are divided into two groups: the medial group, consisting of thick, more myelinated fibers, and the lateral group, formed by thin, less myelinated fibers.
The medial group of dorsal root fibers is sent to the posterior cord of the white matter, where each fiber divides in a T-shape into ascending and descending branches. The ascending branches, following upward, come into contact with the cells of the gray matter of the spinal cord in the gelatinous substance and in the dorsal horn, and some of them reach the medulla oblongata, forming thin and wedge-shaped bundles, fasciculi gracilis et cuneatus (see Fig. 880, 881, 882), spinal cord.
The descending branches of the fibers are directed downward and come into contact with the cells of the gray matter of the posterior columns over six to seven underlying segments. Some of these fibers form a bundle in the thoracic and cervical sections of the spinal cord, which has the appearance of a comma on the cross section of the spinal cord and is located between the wedge-shaped and thin bundles; in the lumbar region - the type of medial cord; in the sacral region - a view of the oval bundle of the posterior cord adjacent to the medial surface of the thin bundle.
The lateral group of fibers of the dorsal root is directed to the marginal zone, and then to the posterior column of the gray matter, where it comes into contact with the cells of the dorsal horn located in it.
The fibers extending from the cells of the spinal cord nuclei are directed upward partly along the lateral cord on their side, and partly pass as part of the white commissure to the opposite side of the spinal cord and are also directed upward in the lateral cord.
The ascending tracts (see Fig. 880, 881, 882), starting in the spinal cord, include the following:
- The posterior spinocerebellar tract, tractus spinocerebellaris dorsalis [posterior], is the direct cerebellar tract, conducts impulses from muscle and tendon receptors to the cerebellum. The bodies of the first neurons lie in the spinal ganglion, the bodies of the second neurons lie throughout the entire length of the spinal cord in the thoracic column (thoracic nucleus) of the dorsal horn. The long processes of the second neurons extend outward; Having reached the posterolateral part of the spinal cord on the same side, they turn upward and rise along the lateral cord of the spinal cord, and then follow the inferior cerebellar peduncle to the cortex of the cerebellar vermis.
- The anterior spinocerebellar tract, tractus spinocerebellaris ventralis [anterior], conducts impulses from muscle and tendon receptors to the cerebellum. The bodies of the first neurons lie in the spinal ganglion, and the bodies of the second neurons - in the medial nucleus of the intermediate zone and send part of their fibers through the white commissure to the lateral cords of the opposite side, and part - to the lateral cords of their own side. These fibers reach the anterior outer sections of the lateral funiculi, located anterior to the posterior spinocerebellar tract. Here the fibers turn upward, go along the spinal cord, and then along the medulla oblongata and, having passed the bridge, along the superior cerebellar peduncles, making a second decussation, they reach the cerebellar vermis.
- The spinoolivary tract, tractus spinoolivaris, originates from the cells of the dorsal horns of the gray matter. The axons of these cells cross and rise near the surface of the spinal cord at the border of the lateral and anterior funiculi, ending in the olive nuclei. The fibers of this pathway carry information from skin, muscle and tendon receptors.
Rice. 952. Conducting pathways of the internal capsule and cerebral peduncles (semi-schematic). Fig. 951. Capsules and the course of pathways through the internal capsule (semi-schematically).
- The anterior and lateral spinothalamic pathways, tractus spinothalamici ventralis [anterior] et lateralis (see Fig. 947), conduct impulses of pain, temperature (lateral pathway) and tactile (anterior pathway) sensitivity. The cell bodies of the first neurons lie in the spinal ganglia. The processes of the second neurons from the cells of the nucleus of the dorsal horn are directed through the white commissure to the anterior and lateral funiculi of the opposite side. Rising upward, the fibers of these pathways pass in the posterior parts of the medulla oblongata, pons and cerebral peduncles and reach the thalamus as part of the spinal lemniscus, lemniscus spinalis. The bodies of the third neurons of these pathways lie in the thalamus, and their processes are directed to the cerebral cortex as part of the central thalamic radiations through the posterior leg of the internal capsule (Fig. 951, 952).
- The spinal reticular tract, tractus spinoreticularis, consists of fibers that pass as part of the spinothalamic tracts, do not intersect and form bilateral projections to all parts of the stem reticular formation.
- The spinal tegmental tract, tractus spinotectalis, together with the spinothalamic tract passes in the lateral cords of the spinal cord and ends in the plate of the roof of the midbrain.
- The thin bundle, fasciculus gracilis, and the wedge-shaped bundle, fasciculus cuneatus (see Fig. 947), conduct impulses from muscles, joints and tactile sensitivity receptors. The bodies of the first neurons of these pathways are localized in the corresponding spinal ganglia. The axons travel as part of the dorsal roots and, having entered the posterior columns of the spinal cord, take an ascending direction, reaching the nuclei of the medulla oblongata.
The thin bundle occupies a medial position and conducts corresponding impulses from the lower extremities and lower parts of the torso - below the 4th thoracic segment.
The wedge-shaped bundle is formed by fibers starting from the cells of all spinal nodes lying above the 4th thoracic segment.
Having reached the medulla oblongata, the fibers of the thin bundle come into contact with the cells of the nucleus of this bundle, which lies in the tubercle of the thin nucleus; the fibers of the sphenoid fasciculus end in the sphenoid tubercle. The cells of both tubercles are the bodies of the second neurons of the described pathways. Their axons - internal arcuate fibers, fibrae arcuatae internae - are directed forward and upward, pass to the opposite side and, forming a decussation of the medial loops (sensitive decussation), decussatio lemniscorum medialium (decussatio sensoria), with fibers of the opposite side, go as part of the medial loop lemniscus medialis.
Having reached the thalamus, these fibers come into contact with its cells - the bodies of the third neurons of the pathway, which send their processes through the internal capsule to the cerebral cortex.
Spinal fibers
The afferent pathways of the spinal cord are formed with the help of axons, or, as they are also called differently, the endings of neurons. Axons are located only in the spinal cord and do not extend beyond it, and also create connections between all segments of the organ. The atomic structure of these fibers is that the length of the axons is quite large and connects to other nerve endings. Nerve signals are transmitted from the receptors to the central nervous system through the afferent pathways of the spinal cord and brain. All nerve fibers located along the entire length of the spinal cord are involved in this process. The signal to the organs is carried out from different parts of the central nervous system and between neurons. The unimpeded passage of signals from the periphery to the central nervous system is achieved using the spinal cord pathways.
How do they work together and how are they different?
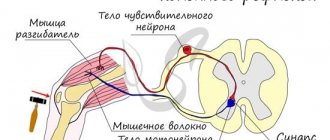
Afferent neurons typically have two axons that carry electrochemical signals to the spinal column or brain. Once there, the signal travels through a network of interneurons and through the efferent neuron. Afferent-efferent pairs of neurons that pass through the spine control reflexes (such as the knee-jerk reaction).
Finished works on a similar topic
Course work Afferent and efferent nerve conductors and their role in psychology 410 ₽ Abstract Afferent and efferent nerve conductors and their role in psychology 240 ₽ Test work Afferent and efferent nerve conductors and their role in psychology 240 ₽
Receive completed work or specialist advice on your educational project Find out the cost
Afferent neurons are designed to respond to various stimuli. For example, an afferent neuron designed to respond to heat detects excess heat and sends an impulse through the central nervous system. The efferent neuron then causes the muscles to contract to move the body away from the heat. The skin has sensory receptors for heat, cold, pleasure, pain and pressure.
Afferent neurons have round and smooth cell bodies, while efferent neurons have satellite bodies. Afferent neurons are found in the peripheral nervous system, and efferent neurons are located in the central nervous system. Axons in afferent neurons move from the ganglia (clusters of nerve cells that contain afferent and efferent neurons) to the spinal cord. The long axon is actually connected to the efferent neuron.
Afferent neurons have a single long myelinated dendrite, whereas efferent neurons have shorter dendrites. The dendrite in an afferent neuron is what is responsible for transmitting nerve impulses from the receptors to the cell body, while in an efferent neuron the impulses travel through the dendrite and exit through the neuromuscular junction that is formed between the effectors and the axon.
Posterior and anterior spinocerebellar tracts
The afferent pathways of the cerebellum are unconscious and originate in the lateral cord of the spinal cord, and from there they carry information about the state of the organs of the musculoskeletal system. The anterior spinocerebellar tract enters the cerebellum through the superior peduncle, and therefore passes through the tegmentum of the medulla oblongata, midbrain and pons. The posterior spinocerebellar tract runs in the medulla oblongata and enters through the inferior peduncle.
These two tracts transmit information from ligaments, joint capsules, muscle receptors, tendons, and periosteum to the cerebellum. They are responsible for maintaining balance and coordinating human movements, so their role in the body is very important.
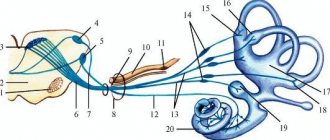
Auditory fibers
This pathway carries information from the receptors of the organ of Corti, which is located in the inner ear. Nerve impulses enter the bridge, which contains the auditory nuclei, along the fibers of the vestibulocochlear nerve. Through the auditory nuclei, information is transmitted to the nuclei of the trapezoid body. After this, the impulses arrive at the subcortical hearing centers, which include the thalamus optic, inferior colliculi and medial geniculate bodies.
In the midbrain, return reactions arise to these auditory stimuli, while the afferent auditory pathways switch to the nuclei of the thalamus, in which auditory stimuli are evaluated - they are responsible for movements that occur involuntarily: walking, running. Auditory radiation begins to emanate from the geniculate bodies - this tract conducts impulses from the internal capsule to the projection center of hearing. Only here does the evaluation of sounds begin to take place. The associative auditory center is located in the posterior part of the temporal gyrus. It is in it that all sounds begin to be perceived as words.
Taste analyzers
The impulses of the afferent pathway of taste analyzers develop from the receptors of the root of the tongue, which are part of the glossopharyngeal nerves and located on the tongue, which are part of the facial nerve. Impulses from them enter the medulla oblongata, and then to the nuclei of the facial and glossopharyngeal nerve. The smallest part of all information received from these impulses is delivered to the cerebellum, thereby forming the nuclear-cerebellar pathway, and provides reflex regulation of the tone of the muscles of the tongue, head and pharynx. Most of the information enters the visual thalamus, after which the impulses reach the uncinus of the temporal lobe, where they are consciously analyzed.
Associative paths, short and long.
Association pathways are the link between the last neuron of the afferent pathway and the first neuron of the efferent pathway.
Associative pathways are chains of interneurons that connect different areas of the cortex within the same hemisphere. In the SM, associative paths connect neighboring segments.
There are short and long associative paths.
Short association fibers connect areas of the cortex of neighboring gyri, without going beyond the lobe of the cerebral hemisphere. These are arcuate fibers located superficially under the bark - at the bottom of the furrows. This also includes fibers that connect cells of neighboring nuclei of the brain stem.
Long association fibers are located under a layer of short association fibers in the cerebral hemispheres. They connect areas of the cortex of different lobes of one hemisphere.
Long associative fibers include: 1. Belt. This is a group of nerve fibers located deep in the vaulted gyrus. Connects areas of the cortex of the frontal, occipital and temporal lobes in the region of the medial surface of the cerebral hemisphere. In ontogenesis, the belt develops earlier than other associative pathways. 2. Upper longitudinal beam. It is localized under the superolateral surface of the cerebral hemisphere, lateral to the corona radiata.
Its fibers connect areas of the cortex of the lower parts of the frontal lobe, the inferior parietal lobule, the temporal and occipital lobes. The formation of the superior longitudinal fasciculus in ontogenesis is associated with the development of the cortical ends of the cutaneous, motor, auditory and visual analyzers.
3. Lower longitudinal fascicle. It is located under the inferomedial surface of the cerebral hemisphere, along the outer wall of the posterior and inferior horns of the lateral ventricle.
Bundles of fibers connect areas of the cortex of the occipital and temporal lobes. The lower longitudinal fasciculus develops earlier than the upper one and provides connection between the cortical end of the visual analyzer and the autonomic centers, and therefore their combined actions. 4. Hook-shaped bundle. Located on the inferolateral surface of the cerebral hemisphere. Connects areas of the cortex of the frontal, temporal and occipital lobes of the hemisphere. Long associative tracts include fibers passing through the stria terminalis, the medullary stria of the thalamus, and the dorsal and medial longitudinal fasciculi.
In the SM, associative fibers are located around the SV in the form of a narrow strip.
These are their own bundles, which are classified as short associative paths. They are part of local reflex arcs, connecting segments of the SC, as well as sensory neurons of the spinal ganglia with motor neurons of the anterior horns of the SC.
In ontogenesis, associative pathways are formed much later compared to commissural and projection pathways.
However, in the future, associative pathways quickly develop and dominate over other pathways.
Commissural pathways. Projection pathways: a) ascending (afferent) fiber systems.
Commissural nerve tracts are pathways of the central nervous system that connect the symmetrical parts of the cerebral hemispheres or other parts of the central nervous system. Commissural (commissural) pathways connect areas of the cortex of the right and left hemispheres of the brain and ensure the unity of brain activity.
Commissural, that is, commissural tracts connect both symmetrical and asymmetrical areas of the cortex of different hemispheres.
The commissural tracts include: corpus callosum, anterior commissure of the brain, posterior commissure of the brain, commissure of the fornix.
Corpus callosum. Among other commissures, it is phylogenetically the youngest formation.
It consists of transversely directed nerve fibers connecting similar areas of the neocortex of the right and left hemispheres of the cerebrum with each other. Part of the fibers that form the beak of the corpus callosum connects both thalami and the heads of the caudate nuclei, ensuring synchronous activity not only of the cortex of the right and left hemispheres, but also of the subcortex. Anterior commissure of the brain. Represented by fibers belonging to the pathways of the olfactory brain.
The anterior commissure of the brain is located in front of the columns of the medullary vault and consists of anterior and posterior parts. The formation of the anterior part of the commissure is associated with the formation of the ancient cortex, and the posterior part is associated with the formation of the new cortex. It is suggested that the anterior commissure is important for the paired activity of not only the olfactory, but also the auditory and visual analyzers. The anterior commissure provides interhemispheric integration of impulses.
Posterior commissure of the brain. Located in the posterior wall of the third ventricle. The fibers that make up the commissure connect the thalami cushions to each other. The commissure of the fornix (hippocampal commissure). This is an ancient formation related to the pathways of the olfactory brain. The commissure fibers of the fornix connect the structures of the hippocampus of the right and left hemispheres and ensure the synchronous functioning of the hippocampus.
The commissure of the fornix is located between the lower surface of the splenium of the corpus callosum and the crura of the fornix.
Projection nerve tracts are pathways of the central nervous system that connect the cerebral cortex with the periphery and pass through various parts of the central nervous system. Projection nerve pathways are divided into afferent (ascending) and efferent (descending).
The ascending pathways of the lateral column include the following: spinothalamic, or tractus spinothalamicus - fibers of the second neurons of the pain and temperature, partly tactile sense, which passed into the opposite lateral column after crossing in the anterior gray commissure.
In addition to the simplest reflex arcs that arise within one segment of the SC, there are also intersegmental reflex ascending and descending pathways.
Let's look at the ascending paths. When performing the knee reflex, the extensor muscle contracts and the leg involuntarily extends. Signals from the sensory neuron, indicating that the leg has changed position, travel up the collateral of the sensory neuron.
This collateral passes in the spinal cord to the thalamus. This is where incoming signals are filtered. For example, if the signal is single and weak, then it simply does not pass through.
Such a system allows you to regulate the passage of signals to the cortex and makes it possible to respond to more important signals with the greatest speed. After passing through the thalamus, the nerve impulse arrives at the neurons of the somatosensory cortex of the cerebral hemispheres of the telencephalon. In this case, the person has the feeling that the leg has straightened.
In order to return it to its original position, the nerve impulse is transmitted to the motor cortex of the cerebral hemispheres, where movement programs are built. Intersegmental ascending pathways allow us to control voluntary movements triggered at different levels of the SC.
Visual analyzers
The afferent pathways of the central nervous system of the visual analyzer begin from the cones and rods of the retina of the eyeball. The impulses enter the optic chiasm as part of the optic nerves, and then along the tract are sent to the subcortical centers of the brain, that is, to the visual thalamus, geniculate lateral bodies and superior colliculus, located in the middle part of the brain.
In the midbrain, a response to these stimuli occurs, and in the nuclei of the thalamus an unconscious assessment of impulses begins that provide involuntary movements reproduced by a person. The main unconscious movements are running and walking. In the projection center of vision or in the calcarine sulcus of the occipital lobe of the brain, impulses arrive along the optic radiation from the geniculate bodies located in the internal capsule, after which a full analysis of the incoming data begins. In the cortex, which is adjacent to the calcarine sulcus, the central part responsible for visual memory, which is also called the associative visual center, finds its location.
Ascending (afferent) pathways starting in the brain stem
The medial lemniscus, trigeminal lemniscus, ascending tract of the auditory analyzer, optic radiation, and thalamic radiation begin in the brain stem.
1. The medial lemniscus as a continuation of the thin and wedge-shaped fascicles was described earlier.
2. The trigeminal loop, lemniscus trigeminalis, is formed by processes of nerve cells that make up the sensory nuclei of the trigeminal nerve (V pair), facial nerve (VII pair), glossopharyngeal nerve (IX pair) and vagus nerve (X pair).
The axons of afferent neurons located in the trigeminal ganglion approach the sensory nuclei of the trigeminal nerve. The common sensory nucleus of the other three nerves - the nucleus of the solitary tract - is approached by axons of afferent neurons located in the genu node (VII pair) and in the upper and lower nodes of the IX and X pairs of nerves. The bodies of the first neurons are localized in the listed nodes, and the bodies of the second neurons of the path along which impulses from the head receptors are transmitted are located in the sensitive nuclei.
The fibers of the trigeminal lemniscus pass to the opposite side (some of the fibers follow on their side) and reach the thalamus, where they end in its nuclei.
The nerve cells of the thalamus are the bodies of the third neurons of the ascending tracts of the cranial nerves, the axons of which, as part of the central thalamic radiates, through the internal capsule are directed to the cerebral cortex (postcentral gyrus).
3. The ascending path of the auditory analyzer has, as its first neurons, cells located in the ganglion of the cochlear part of the vestibulocochlear nerve. The axons of these cells approach the cells of the anterior and posterior cochlear nuclei (second neurons). The processes of the second neurons, moving to the opposite side, form a trapezoidal body, and then take an ascending direction and are called the lateral loop, lemniscus lateralis. These fibers end on the bodies of the third neurons of the auditory pathway, located in the lateral geniculate body. The processes of the third neurons form the auditory radiation, radiatio acustica, which runs from the medial geniculate body through the posterior limb of the internal capsule to the middle part of the superior temporal gyrus.
4. Visual radiance, radiatio optica (see Fig. 951), connects the subcortical centers of vision with the cortex of the calcarine sulcus.
The optic radiance includes two systems of ascending fibers:
- the geniculate-cortical optic tract, which originates from the cells of the lateral geniculate body;
- the cushion-cortical tract, starting from the cells of the nucleus located in the cushion of the thalamus; in humans it is poorly developed.
The set of these fibers is designated as posterior thalamic radiation, radiationes thalamicae posteriores.
Ascending to the cerebral cortex, both systems pass through the posterior limb of the internal capsule.
5. Thalamic radiations, radiationes thalamicae (see Fig. 947), are formed by processes of thalamic cells and constitute the final sections of the ascending tracts of the cortical direction.
The thalamic radiations include:
- anterior thalamic radiations, radiationes thalamicae anteriores, are radial fibers of the white matter of the cerebral hemispheres. They begin from the superior medial nucleus of the thalamus and are directed through the anterior limb of the internal capsule to the cortex of the lateral and inferior surfaces of the frontal lobe. Some of the fibers of the anterior thalamic radiates connect the anterior group of thalamic nuclei with the cortex of the medial surface of the frontal lobes and the anterior part of the cingulate gyrus;
- central thalamic radiations, radiationes thalamicae centrales, are radial fibers connecting the ventrolateral group of thalamic nuclei with the cortex of the pre- and postcentral gyrus, as well as with the adjacent sections of the cortex of the frontal and parietal lobes. They pass as part of the posterior limb of the internal capsule;
- the lower peduncle of the thalamus, pedunculus thalami inferior, contains radial fibers connecting the thalamic cushion and medial geniculate bodies with areas of the temporal choir;
- posterior thalamic radiates (see earlier).
Olfactory analyzer
The afferent pathway of the olfactory analyzer originates from the receptors of the mucous membrane, localized in the upper part of the nasal passage. After this, the impulses are sent to the axons of the olfactory bulbs, and they flow along the fibers of the olfactory nerves. Then the impulses are sent to the projection center of smell, which is located in the area of the parahippocampal gyrus and uncus. These impulses follow along the tract to the cortex of the temporal lobe of the brain. Most of the information received from the olfactory receptors is sent to the subcortical centers, which are located in the middle and intermediate parts of the brain. The subcortical centers of the brain, in response to olfactory stimuli, provide reflex regulation of muscle tone.
Based on this, it can be determined that the main feature of olfactory receptors is that nerve impulses initially enter the cortex of the cerebral hemispheres, and not the subcortical centers of smell. In this regard, a person first smells the smell, then begins to evaluate it, and only after that an unconscious coloring of the stimulus is formed in the brain on an emotional level. The entire process takes only a fraction of a second.
Intraoperative identification of functional areas of the cortex and brain pathways
Back in the 30s of the last century, the first steps were taken in determining the localization of the cortical functional organization during neurosurgical operations [46, 76].
Initially, only direct stimulation of the cortex was used during the removal of tumors and epileptogenic zones in conscious patients, which made it possible to assess the relationships between consciousness, motor acts and language production at the stages of surgical intervention. Later, in the 70s of the 20th century, monitoring of somatosensory evoked potentials (SSEPs) entered neurophysiological practice [73], and more recently, in the 90s, motor evoked potentials appeared, which made it possible to more accurately localize the functional areas of the cerebral cortex and pathways . In 1978, the phenomenon of phase reversal of the cortical component of SSEP in the region of the central gyri and the identification of the central sulcus was described [51], which has found wide application in the surgery of intracerebral tumors [28, 59, 63]. In 1937, W. Penfield [76] described a technique for direct stimulation of the cortex using rhythmic (50-60 Hz) bipolar impulses. This approach is mainly used to map the sensory and motor areas of speech in conscious patients, although it is also used in patients under general anesthesia [63]. Also currently used is the technique of stimulation in bursts (trains) of 4-5 pulses [90] for mapping and long-term monitoring of cortical-subcortical conductors [17].
Arousal surgery and direct electrical stimulation of the cortex and pathways are the “gold standard” for resection of gliomas located near speech areas and other functionally significant structures [36, 38, 42, 74]. During such operations, functional brain mapping is carried out by a neurosurgeon together with a neuropsychologist and neurophysiologist, the latter adjusting stimulation parameters while simultaneously monitoring the state of spontaneous cortical activity using an electrocorticogram (ECoG) to detect epileptic activity. This approach makes it possible to prevent the development of convulsive syndrome during prolonged rhythmic electrical stimulation. At the first stage, electrical stimulation reveals the localization of cortical zones of language functions, with each functionally important zone being marked [2, 11, 12].
The stimulation zone is considered functionally significant when speech disturbances are observed three times in a row after subsequent stimuli, and after the end of stimulation, restoration of speech (language) function is noted. The type of speech impairment is verified by a neuropsychologist, and the severity of speech impairment is assessed using the National Institutes of Health Stroke Scale [92]. The next stage is tumor resection taking into account mapping data, with periodic subcortical stimulation performed to search for functionally significant white matter pathways [2].
Thus, the use of intraoperative mapping and electrophysiological monitoring allows surgeons to remove the maximum volume of tumor with minimal impairment of neurological functions [31, 54, 61].
4. Neuropsychological methods of intraoperative research
The study of cognitive functions in dynamics (before and after surgery) is carried out using a well-selected set of neuropsychological tests, which must satisfy several criteria: 1) allow for the assessment of a number of cognitive functions, while the tests must be sensitive enough to determine the effect of the tumor and the treatment performed; 2) the testing procedure must be standardized to allow comparison of the results of a dynamic examination of patients; 3) the presence of standards for conducting tests will make it possible to objectively assess the performance of tests by patients; 4) tests must be sufficiently reliable and insensitive to repeated testing to obtain accurate data during a dynamic examination; 5) tests should have alternative forms for retesting; 6) to prevent fatigue, the total testing time should not exceed 30-40 minutes.
A set of neuropsychological testing is selected individually for each patient. Since tumors often impair memory, attention, speed of information processing, executive functions and speech, appropriate tests must be used during testing. Examples include tests for verbal associations (associations for a given letter of the alphabet and for a certain semantic category), finding similarities between two concepts (for example, “what is common between an apple and a banana”), a test of following route A and B (allows you to evaluate visual attention, distribution and switching of attention) [65], memorizing a list of words (allows you to assess auditory-verbal memory) [55, 60, 83], encryption test (allows you to assess the speed of information processing) [93], test for fine motor skills [53]. Depending on the location of the lesion, the study can be supplemented with the Wechsler Intelligence Test (WAIS-IV), which provides a summary assessment of verbal and nonverbal intelligence [93]. The Rey Complex Figure Test assesses visual constructive functions. Tests for memorizing non-verbalizable figures (Rey Visual Design Learning Test, Rey Complex Figure Test) [65, 89], tests for speech and non-speech working memory (Digit Span Forward, Digit Span Backward, Spatial Span Forward, Spatial Span Backward) are also used [ 65, 89], tests for speech research [22].
Intraoperative mapping uses a small set of relatively simple tests that can be performed during intraoperative awakening. Each test must include a sufficient number of samples to allow continuous testing of function over the course of brain stimulation. As an example, we present several tests used for intraoperative localization of motor and speech areas, most often determined during such operations. To localize the motor zone of the hand, a test for clenching and unclenching a fist is used; to localize the motor zone of the leg, a test for flexing and extending the leg at the knee joint is used. To assess speech functions, a picture naming test is used, and naming verbs is more sensitive to localizing Broca's area than naming nouns. Ordinal counting in forward and reverse order allows you to evaluate the fluency of speech and switching.
Thus, neuropsychological examination of cognitive functions is an important component of a comprehensive examination of patients with brain tumors in the pre- and postoperative period, determining both surgical tactics and subsequent rehabilitation measures.
Vestibular tract
The vestibular afferent pathway begins from the receptors of the semicircular canal of the inner ear, the uterus and the receptors that make up this organ. This tract in the central nervous system is responsible for coordinating movements and maintaining balance during physical and vestibular stress.
Afferent centripetal pathways and the peculiarity of their structure indicate that a person needs to make a lot of effort to maintain the health and integrity of each organ individually and together. Each component of this pathway provides the body with all the necessary information, helps to immediately process it and carry out all vital processes. This is important in the functioning of the entire organism as a whole and individual organs.