Nervous system
Types of nervous systems
There are several types of organization of the nervous system, represented in various systematic groups of animals.
- Diffuse nervous system
- presented in coelenterates. Nerve cells form a diffuse nerve plexus in the ectoderm throughout the animal's body, and when one part of the plexus is strongly stimulated, a generalized response occurs - the whole body reacts. - Stem nervous system
(
orthogon
) - some nerve cells are collected into
nerve trunks
, along with which the diffuse subcutaneous plexus is preserved. This type of nervous system is present in flatworms and nematodes (in the latter the diffuse plexus is greatly reduced), as well as many other groups of protostomes - for example, gastrotrichs and cephalopods. - Nodal nervous system
, or complex ganglion system, is represented in annelids, arthropods, mollusks and other groups of invertebrates.
Most of the cells of the central nervous system are collected in nerve nodes - ganglia. In many animals, the cells are specialized and serve individual organs. In some molluscs (for example, cephalopods) and arthropods, a complex association of specialized ganglia with developed connections between them arises - a single brain or cephalothoracic nerve mass (in spiders). of the protocerebrum
have a particularly complex structure . - A tubular nervous system (neural tube) is characteristic of chordates.
Nervous system of various animals
Nervous system of cnidarians and ctenophores
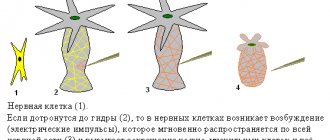
Cnidarians are considered the most primitive animals that have a nervous system. In polyps, it represents a primitive subepithelial nervous network ( nervous plexus
), entwining the entire body of the animal and consisting of neurons of different types (sensitive and ganglion cells), connected to each other by processes (
diffuse nervous system
), their especially dense plexuses are formed on the oral and aboral poles of the body.
Irritation causes rapid conduction of excitation through the body of the hydra and leads to contraction of the entire body, due to the contraction of epithelial-muscle cells of the ectoderm and at the same time their relaxation in the endoderm. Jellyfish are more complex than polyps; a central section begins to separate in their nervous system. In addition to the subcutaneous nerve plexus, they have ganglia along the edge of the umbrella, connected by processes of nerve cells into a nerve ring
, from which the muscle fibers of the velum and
rhopalia
- structures containing various sensory organs (
diffuse nodular nervous system
). Greater centralization is observed in scyphojellyfish and especially box jellyfish; 8 ganglia, corresponding to 8 rhopalia, reach quite large sizes.
The nervous system of ctenophores includes a subepithelial nerve plexus with condensations along rows of paddle plates that converge to the base of a complex aboral sensory organ. In some ctenophores, nearby nerve ganglia have been described.
Nervous system of protostomes
Flatworms
have a nervous system already divided into central and peripheral sections.
In general, the nervous system resembles a regular lattice - this type of structure was called orthogon
.
It consists of a cerebral ganglion, which in many groups surrounds the statocysts (endonal medulla), which is connected to the nerve trunks
running along the body and connected by annular transverse bridges (
commissures
).
Nerve trunks consist of nerve fibers extending from nerve cells scattered along their course. In some groups, the nervous system is quite primitive and close to diffuse. The following trends are observed among flatworms: ordering of the subcutaneous plexus with separation of trunks and commissures, an increase in the size of the cerebral ganglion, which turns into the central control apparatus, immersion of the nervous system into the thickness of the body; and, finally, a decrease in the number of nerve trunks (in some groups only two abdominal (lateral) trunks
).
In nemerteans, the central part of the nervous system is represented by a pair of connected double ganglia, located above and below the proboscis sheath, connected by commissures and reaching a significant size. Nerve trunks go back from the ganglia, usually in pairs, and they are located on the sides of the body. They are also connected by commissures; they are located in the skin-muscle sac or in the parenchyma. Numerous nerves depart from the head node, the most strongly developed are the spinal nerve (often double), abdominal and pharyngeal.
Gastrociliary worms have a suprapharyngeal ganglion, a peripharyngeal nerve ring, and two superficial lateral longitudinal trunks connected by commissures.
Nematodes have a peripharyngeal nerve ring, from which 6 nerve trunks extend forward and backward, the largest - the ventral and dorsal trunks - stretch along the corresponding hypodermal ridges. The nerve trunks are connected to each other by semicircular jumpers; they innervate the muscles of the abdominal and dorsal lateral bands, respectively. Nervous system of the nematode Caenorhabditis elegans
has been mapped at the cellular level[1]. Each neuron has been recorded, its origin has been traced, and most, if not all, neural connections are known[2]. In this species, the nervous system is sexually dimorphic: the male and hermaphroditic nervous systems have different numbers of neurons and groups of neurons to perform sex-specific functions.
In Kinorhynchus, the nervous system consists of a peripharyngeal nerve ring and a ventral (abdominal) trunk, on which, in accordance with their inherent body segmentation, ganglion cells are located in groups.
The nervous system of hairworms and priapulids has a similar structure, but their ventral nerve trunk is devoid of thickenings.
Rotifers have a large suprapharyngeal ganglion, from which nerves arise, especially large ones - two nerves that run through the entire body on the sides of the intestine. Smaller ganglia lie in the leg (pedal ganglion) and next to the masseteric stomach ( mastax
).
In acanthocephalans, the nervous system is very simple: inside the proboscis vagina there is an unpaired ganglion, from which thin branches extend forward to the proboscis and two thicker lateral trunks back; they emerge from the proboscis vagina, cross the body cavity, and then go back along its walls.
Annelids have a paired suprapharyngeal ganglion, with peripharyngeal connectives
(connectives, unlike commissures, connect opposite ganglia) connected to the ventral part of the nervous system.
In primitive polychaetes, it consists of two longitudinal nerve cords in which nerve cells are located. In more highly organized forms, they form paired ganglia in each segment of the body ( nervous scala
), and the nerve trunks come closer together.
In the majority of polychaetes, the paired ganglia merge ( ventral nerve cord
); in some, their connectives also merge.
Numerous nerves depart from the ganglia to the organs of their segment. In the series of polychaetes, the nervous system is immersed from under the epithelium into the thickness of the muscles or even under the skin-muscular sac. Ganglia of different segments can be concentrated if their segments merge. Similar trends are observed in oligochaetes. In leeches, the nerve chain lying in the abdominal lacunar canal consists of 20 or more ganglia, and the first 4 ganglia ( subpharyngeal ganglion
) and the last 7 are combined into one.
In echiurids, the nervous system is poorly developed - the peripharyngeal nerve ring is connected to the abdominal trunk, but the nerve cells are scattered evenly throughout them and do not form nodes anywhere.
Sipunculids have a suprapharyngeal nerve ganglion, a peripharyngeal nerve ring, and a nerveless ventral trunk lying on the inside of the body cavity.
Tardigrades have a suprapharyngeal ganglion, peripharyngeal connectives, and a ventral chain with 5 paired ganglia.
Onychophorans have a primitive nervous system. The brain consists of three sections: the protocerebrum innervates the eyes, the deutocerebrum innervates the antennae, and the tritocerebrum innervates the foregut. Nerves extend from the peripharyngeal connectives to the jaws and oral papillae, and the connectives themselves pass into distant abdominal trunks, evenly covered with nerve cells and connected by thin commissures.
Nervous system of arthropods
In arthropods, the nervous system is composed of a paired suprapharyngeal ganglion, consisting of several connected nerve ganglia (brain), peripharyngeal connectives and a ventral nerve cord, consisting of two parallel trunks. In most groups, the brain is divided into three sections - proto-
,
deutocerebrum
and
tritocerebrum
. Each body segment has a pair of nerve ganglia, but the fusion of ganglia to form large nerve centers is often observed; for example, the subpharyngeal ganglion consists of several pairs of fused ganglia - it controls the salivary glands and some muscles of the esophagus.
In a number of crustaceans, in general, the same trends are observed as in annelids: the convergence of a pair of abdominal nerve trunks, the fusion of paired nodes of one body segment (that is, the formation of the abdominal nerve chain), the fusion of its nodes in the longitudinal direction as the body segments unite. Thus, crabs have only two nerve masses - the brain and a nerve mass in the chest, while copepods and barnacles form a single compact formation, penetrated by the canal of the digestive system. The brain of crayfish consists of paired lobes - the protocerebrum, from which the optic nerves, which have ganglion clusters of nerve cells, depart, and the deutocerebrum, which innervates antennae I. Usually, a tritocerebrum is also added, formed by the fused nodes of the antennal segment II, the nerves to which usually arise from the peripharyngeal connectives. Crustaceans have a developed sympathetic nervous system
, consisting of a medulla and an unpaired
sympathetic nerve
, which has several ganglia and innervates the intestines.
An important role in the physiology of crayfish is played by neurosecretory cells
located in various parts of the nervous system and secreting
neurohormones
.
The brain of centipedes has a complex structure, most likely formed by many ganglia. The subpharyngeal ganglion innervates all oral limbs, from which a long paired longitudinal nerve trunk begins, on which there is one paired ganglion in each segment (in bipedal centipedes, in each segment, starting from the fifth, there are two pairs of ganglia located one after the other).
The nervous system of insects, also consisting of the brain and the ventral nerve cord, can achieve significant development and specialization of individual elements. The brain consists of three typical sections, each of which consists of several ganglia separated by layers of nerve fibers. Important associative
protocerebrum. Social insects (ants, bees, termites) have especially developed brains. The abdominal nerve chain consists of the subpharyngeal ganglion, which innervates the oral limbs, three large thoracic ganglia and abdominal ganglia (no more than 11). In most species, more than 8 ganglia are not found in adulthood; in many, these also merge, giving rise to large ganglion masses. It can go so far as to form only one ganglion mass in the thorax, innervating both the thorax and the abdomen of the insect (for example, in some flies). During ontogenesis, ganglia often unite. Sympathetic nerves arise from the brain. Almost all parts of the nervous system contain neurosecretory cells.
In horseshoe crabs, the brain is not externally divided, but has a complex histological structure. Thickened peripharyngeal connectives innervate the chelicerae, all limbs of the cephalothorax and gill covers. The abdominal nerve cord consists of 6 ganglia, the posterior one is formed by the fusion of several. The nerves of the abdominal limbs are connected by longitudinal lateral trunks.
The nervous system of arachnids has a clear tendency to concentrate. The brain consists only of protocerebrum and tritocerebrum due to the lack of structures innervated by deutocerebrum. The metamerism of the ventral nerve chain is most clearly preserved in scorpions - they have a large ganglion mass in the chest and 7 ganglia in the abdomen, in salpugs there is only 1, and in spiders all the ganglia have merged into the cephalothorax nerve mass; in harvestmen and ticks there is no distinction between it and the brain.
Sea spiders, like all chelicerates, do not have a deuterocerebrum. The ventral nerve cord in different species contains from 4-5 ganglia to one continuous ganglion mass.
Nervous system of mollusks
In primitive chiton mollusks, the nervous system consists of a peripharyngeal ring (innervates the head) and 4 longitudinal trunks - two pedal
(innervate the “leg”), which are connected in no particular order by numerous commissures, and two
pleurovisceral
, which are located outward and above the pedal ones (innervate the visceral sac, and connect above the powder). The pedal and pleurovisceral trunks on one side are also connected by many jumpers.
The nervous system of monoplacophorans is structured similarly, but their pedal trunks are connected by only one bridge.
In more developed forms, as a result of the concentration of nerve cells, several pairs of ganglia are formed, which are shifted to the anterior end of the body, with the suprapharyngeal node (brain) receiving the greatest development.
Nervous system in the tentacle clade
Phoronids have primitive subcutaneous plexuses of a diffuse type, and only at the anterior end of the body, between the oral and anal openings, do they condense into a primitive “ganglion” [3]. Unlike other tentacles, the primitiveness of the nervous system of the phoronodes is not the result of degeneration, but is primary.
In bryozoans, due to a sedentary lifestyle, the nervous system is greatly simplified: at the anterior end of the body, between the oral and anal openings, there is a “ganglion”, from which nerves extend to the tentacles and all organs of the bryozoan; The colony does not have a common nervous system. The sense organ is the lophophore[4].
In brachiopods, the nervous system is simplified due to a sedentary lifestyle. It consists of a peripharyngeal nerve ring, with nerve trunks extending from it into the mantle and lophophore. At the anterior edge of the mantle, some species have pigmented areas that react to light; also, the marginal cilia of the mantle are sensitive to touch. It is curious that brachiopod larvae have eyes and statocysts, which are lost during development during the transition to a sessile lifestyle [5].
Nervous system of deuterostomes
| This section of the article has not been written. According to the plan of one or more Wikipedia contributors, a special section should be located in this place. You can help by writing this section. This mark was set on May 17, 2021 . |
Nervous system of echinoderms
The nervous system of echinoderms is primitive, consisting of three separate subsystems, built, like the body of echinoderms, on a radial plan: the nerve ring and radial nerve cords. In the oral wall of the body there are two nervous systems - sensory and motor. In the aboral wall of the body there is only motor activity. The sense organs of echinoderms are quite diverse, but primitive in structure. They are diffusely distributed throughout the body in the form of various sensitive cells (functions of touch, chemical sense, vision). Light-sensitive cells may be collected in the ocelli. In sea stars, the eyes are located at the ends of the rays, and in sea urchins - around the anus.
Nervous system of tunicates
In the larval - free-swimming - state they have a developed nervous system - a neural tube with a pronounced brain and spinal cord, a pair of eyes and a statocyst; but with the transition to the adult stage, due to the latter’s usually immobile lifestyle, the nervous system is significantly reduced: for example, to a single primitive ganglion (without an internal cavity) in ascidians. The degree of reduction is proportional to the level of immobility of the adult: the greatest in sessile ascidians, less pronounced in mobile salps, reduction does not occur only in appendiculars, which lead an active free-swimming lifestyle, close to that of the larvae.
Vertebrate nervous system
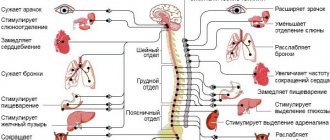
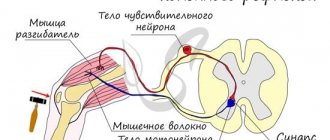
The nervous system of vertebrates is often divided into the central nervous system (CNS) and the peripheral nervous system (PNS). The central nervous system consists of the brain and spinal cord. The PNS is made up of other nerves and neurons that do not lie within the CNS. The vast majority of nerves (which are actually the axons of neurons) belong to the PNS. The peripheral nervous system is divided into the somatic nervous system and the autonomic nervous system.
The somatic nervous system is responsible for coordinating body movement and receiving and transmitting external stimuli. This system regulates actions that are under conscious control.
The autonomic nervous system is divided into parasympathetic and sympathetic parts. The sympathetic nervous system is responsible for responses to danger or stress, and, among many physiological changes, can cause an increase in heart rate and blood pressure and arousal of the senses due to an increase in adrenaline in the blood. The parasympathetic nervous system, on the other hand, is responsible for the state of rest, and ensures that the pupil contracts, slows the heart, dilates blood vessels and stimulates the digestive and genitourinary systems.
Mammalian nervous system
The nervous system functions as an integral unit with sensory organs, such as the eyes, and is controlled in mammals by the brain. The largest part of the latter is called the cerebral hemispheres (in the occipital region of the skull there are two smaller hemispheres of the cerebellum). The brain connects to the spinal cord. In all mammals, with the exception of monotremes and marsupials, unlike other vertebrates, the right and left cerebral hemispheres are connected to each other by a compact bundle of nerve fibers called the corpus callosum. In the brains of monotremes and marsupials there is no corpus callosum, but the corresponding areas of the hemispheres are also connected by nerve bundles; for example, the anterior commissure connects the right and left olfactory areas with each other. The spinal cord, the main nerve trunk of the body, passes through a canal formed by the foramina of the vertebrae and extends from the brain to the lumbar or sacral spine, depending on the species. On each side of the spinal cord, nerves extend symmetrically to different parts of the body. The sense of touch is, in general terms, provided by certain nerve fibers, countless endings of which are located in the skin. This system is usually supplemented by hairs that act as levers to press on areas riddled with nerves.
Morphological division
The nervous system of mammals and humans is divided according to morphological characteristics into central (brain and spinal cord) and peripheral (composed of nerves extending from the brain and spinal cord[6]).
The composition of the central nervous system can be represented as follows:
| |||||||||||||||||||
The peripheral nervous system includes cranial nerves, spinal nerves and nerve plexuses
Functional division
- Somatic (animal) nervous system
- Autonomic (autonomic) nervous system Sympathetic division of the autonomic nervous system
- Parasympathetic division of the autonomic nervous system
- Metasympathetic division of the autonomic nervous system (enteric nervous system)
Ontogenesis
Models
At the moment, there is no single position on the development of the nervous system in ontogenesis. The main problem is to assess the level of determinism (predestination) in the development of tissues from germ cells. The most promising models are the mosaic model
and
regulatory model
. Neither one nor the other can fully explain the development of the nervous system.
- The mosaic model assumes complete determination of the fate of an individual cell throughout ontogeny.
- The regulatory model assumes the random and variable development of individual cells, with only the neural direction being deterministic (that is, any cell of a certain group of cells can become anything within the scope of development for this group of cells).
For invertebrates, the mosaic model is almost flawless—the degree of determination of their blastomeres is very high. But for vertebrates everything is much more complicated. A certain role of determination here is undoubted. Already at the sixteen-cell stage of development of the vertebrate blastula, it is possible to say with a fair degree of certainty which blastomere is not
the predecessor of a certain organ.
Marcus Jacobson
in 1985 he introduced a clonal model of brain development (close to regulatory). He suggested that the fate of individual groups of cells representing the progeny of an individual blastomere, that is, “clones” of this blastomere, is determined. Moody and Takasaki (independently) developed this model in 1987. A map of the 32-cell blastula stage was constructed. For example, it has been established that descendants of the D2 blastomere (vegetative pole) are always found in the medulla oblongata. On the other hand, the descendants of almost all blastomeres of the animal pole do not have pronounced determination. In different organisms of the same species, they may or may not occur in certain parts of the brain.
Regulatory Mechanisms
It was found that the development of each blastomere depends on the presence and concentration of specific substances - paracrine factors, which are secreted by other blastomeres. For example, in an in vitro
with the apical part of the blastula, it turned out that in the absence of activin (paracrine factor of the vegetative pole), the cells develop into ordinary epidermis, and in its presence, depending on the concentration, in increasing order: mesenchymal cells, smooth muscle cells, notochord cells or cardiac muscle cells.
All substances that determine the behavior and fate of the cells that perceive them, depending on the dose (concentration) of the substance in a given area of the multicellular embryo, are called morphogens
.
Some cells secrete soluble active molecules (morphogens) into the extracellular space, decreasing from their source along a concentration gradient.
That group of cells whose location and purpose is specified within the same boundaries (with the help of morphogens) is called a morphogenetic field
. The fate of the morphogenetic field itself is strictly determined. Each specific morphogenetic field is responsible for the formation of a specific organ, even if this group of cells is transplanted into different parts of the embryo. The fates of individual cells within the field are not so rigidly fixed, so that they can, within certain limits, change their purpose, replenishing the functions of cells lost by the field. The concept of the morphogenetic field is a more general concept; in relation to the nervous system, it corresponds to the regulatory model.
The concept of embryonic induction is closely related to the concepts of morphogen and morphogenetic field. This phenomenon, also common to all body systems, was first shown in the development of the neural tube.
Development of the vertebrate nervous system
The nervous system is formed from the ectoderm, the outermost of the three germ layers. A paracrine interaction begins between the cells of the mesoderm and ectoderm, that is, a special substance is produced in the mesoderm - neuronal growth factor, which is transferred to the ectoderm. Under the influence of neuronal growth factor, part of the ectodermal cells turns into neuroepithelial cells, and the formation of neuroepithelial cells occurs very quickly - at a rate of 250,000 pieces per minute. This process is called neuronal induction (a special case of embryonic induction).
As a result, a neural plate is formed, which consists of identical cells. From it the neural folds are formed, and from them the neural tube, which is separated from the ectoderm (it is the change in the types of cadherins, cell adhesion molecules, that is responsible for the formation of the neural tube and neural crest), going under it. The mechanism of neurulation differs somewhat between lower and higher vertebrates. The neural tube does not close along its entire length simultaneously. First of all, the closure occurs in the middle part, then this process spreads to its rear and front ends. At the ends of the tube, two open sections remain - the anterior and posterior neuropores.
Then the process of differentiation of neuroepithelial cells into neuroblasts and glioblasts occurs. Glioblasts give rise to astrocytes, oligodendrocytes and epindymal cells. Neuroblasts become neurons. Next, the process of migration occurs - neurons move to where they will perform their function. Due to the growth cone, the neuron crawls like an amoeba, and the processes of glial cells indicate its path. The next stage is aggregation (sticking together of neurons of the same type, for example, those involved in the formation of the cerebellum, thalamus, etc.). Neurons recognize each other thanks to surface ligands - special molecules found on their membranes. Having united, the neurons are arranged in the order necessary for a given structure.
After this, the nervous system matures. An axon grows from the growth cone of the neuron, and dendrites grow from the body.
Then fasciculation occurs - the union of similar axons (formation of nerves).
The last stage is the programmed death of those nerve cells in which a malfunction occurred during the formation of the nervous system (about 8% of cells send their axon to the wrong place).