Introduction
The pathways of the nervous system and the complex reflex arcs consisting of them are the most important and complex section of neurology. It is important because it affirms the cellular nature of the nervous system (neural doctrine) and shows the ordered nature of the arrangement and connections of neurons (in the form of reflex arcs), which underlies its regulatory function.
There is a significant difference from the method of descriptive anatomy. The latter makes it possible to demonstrate the shape, size and localization of a particular formation of the nervous system, as well as its belonging to gray or white matter, but does not reveal at all the structural organization of the nervous system and the mechanisms of its functioning.
This “separation” of structure from function, which is dangerous for the worldview, eliminates the systematic approach to the nervous system in the form of the study of reflex arcs. Here the emphasis is placed precisely on the presence of connections between neurons, on their interaction, leading to the functioning of both the nervous system itself and the entire organism. However, at the same time, the number of mental operations among students increases (synthesis is added to analysis), which increases the complexity of mastering the material and its subjective complexity. Nevertheless, only the study of the nervous system as a set of reflex arcs allows us to understand its organization and functional significance. Finally, only knowledge of neural connections and interactions allows for topical diagnosis of damage to the nervous system, i.e. take a meaningful approach to the diagnosis and treatment of nervous and many other diseases and injuries.
How do the concepts of “conducting path” and “reflex arc” relate to each other? Here it should be clearly understood that any conductive path is part of one or another reflex arc. Since there are two main links in the reflex arc: afferent and efferent, the pathways are classified into afferent and efferent. Taking into account the hierarchical principle of construction of the central nervous system (the presence of higher and lower nerve centers subordinate to them) and the possibility of closing reflex arcs at the level of higher nerve centers, it is clear that both afferent and efferent pathways should be localized in both the peripheral and central parts nervous system. Since the closure of somatic reflex arcs (the connection of afferent and efferent links through interneurons) always occurs in the central nervous system, the latter also contain an associative link and corresponding associative pathways, localized only within the central nervous system.
Afferent nerve pathways conduct impulses from the receptor to the nerve center and are sensitive. Afferent nerve pathways ending in the projection centers of the cerebral cortex are classified as pathways of conscious sensitivity. The same afferent pathways that end in the subcortical sensory nerve centers are classified as pathways of unconscious sensitivity.
Efferent nerve pathways conduct impulses from nerve centers to the working organ. Since we are talking here only about the somatic nervous system, the working organ is the skeletal muscle, therefore the efferent nerve pathways are called motor. Depending on which nerve centers the efferent pathways are connected to, the latter are responsible for performing both conscious and unconscious movements.
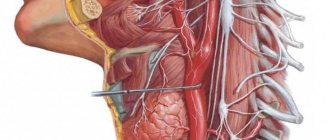
Any conducting pathway (afferent, associative or efferent), depending on the level of closure and complexity of the reflex arc, can be single-neuron or multi-neuron (several neurons connected in series in a chain). If we consider a multineuron pathway as a chain, then within its boundaries we can distinguish links represented by the corresponding neurons. Compactly located neuron bodies form nerve centers (nodal, nuclear or screen type), and axons collected in bundles form nerve tracts. Thus, a multineural pathway consists of nerve centers and tracts. In this case, the nerve centers and tracts of the same pathway are localized in certain but different parts of the nervous system. Each tract within the central nervous system conducts nerve impulses usually in one direction and in most cases - of the same functional content. It should be clearly understood how the tracts within the central nervous system differ from the bundles of fibers that form the cranial or spinal nerves. Nerves contain both afferent and efferent fibers, and different afferent fibers can carry different sensory impulses.
In the future, material will be presented that concerns primarily the somatic part of the nervous system.
DESCENDING PROJECTION PATHWAYS
General characteristics of motor descending pathways:
1.2-neuron structure scheme;
2.the fibers of 1 neuron cross over;
3.2 neuron - in the anterior horns of the spinal cord.
Descending projection pathways (effector, efferent) conduct impulses from the cortex, subcortical centers to the underlying sections, to the nuclei of the brain stem and the motor nuclei of the anterior horns of the spinal cord. These pathways can be divided into two groups: 1) the main motor, or pyramidal, pathway (cortical-nuclear and corticospinal tracts) carries impulses of voluntary movements from the cerebral cortex to the skeletal muscles of the head, neck, trunk, and limbs through the corresponding motor nuclei of the brain and spinal cord; 2) extrapyramidal motor pathways transmit impulses from the subcortical centers to the motor nuclei of the cranial and spinal nerves, and then to the muscles.
The pyramidal tract includes a system of fibers along which motor impulses from the cerebral cortex, from the precentral gyrus, from gigantopyramidal neurons (Betz cells) are sent to the motor nuclei of the cranial nerves and the anterior horns of the spinal cord, and from them to the skeletal muscles. Taking into account the direction of the fibers, as well as the location of the bundles in the brain stem and cords of the spinal cord, the pyramidal tract is divided into three parts: 1) corticonuclear - to the nuclei of the cranial nerves; 2) lateral corticospinal (pyramidal ) - to the nuclei of the anterior horns of the spinal cord; 3) anterior corticospinal (pyramidal) - also to the anterior horns of the spinal cord.
The corticonuclear tract is a bundle of processes of gigantopyramidal neurons, which from the cortex of the lower third of the precentral gyrus descend to the internal capsule and pass through its genu. Next, the fibers of the cortical-nuclear tract go to the base of the cerebral peduncle. Starting from the midbrain and further, in the pons and medulla oblongata, the fibers of the cortical-nuclear tract pass to the opposite side to the motor nuclei of cranial nerves III and IV - in the midbrain, V, VI, VII - in the pons, IX, X, XI, XII - in the medulla oblongata. In these nuclei the corticonuclear (pyramidal) pathway ends. Its constituent fibers form synapses with the motor cells of these nuclei. The processes of the mentioned motor cells leave the brain as part of the corresponding cranial nerves and are directed to the skeletal muscles of the head and neck and innervate them.
The lateral and anterior corticospinal (pyramidal) tracts also begin from the giant pyramidal neurons of the precentral gyrus, its upper 2/3 . The axons of these cells are directed to the internal capsule, pass through the anterior part of its posterior peduncle (immediately behind the fibers of the corticonuclear tract), and descend to the base of the cerebral peduncle. Further, the corticospinal fibers descending, pierce the bundles of pons fibers running in the transverse direction and exit into the medulla oblongata , where on its anterior (lower) surface they form protruding ridges - pyramids . In the lower part of the medulla oblongata, part of the fibers passes to the opposite side and continues in the lateral funiculus of the spinal cord, gradually ending in the anterior horns of the spinal cord with synapses on the motor cells of its nuclei.
This part of the pyramidal tracts, involved in the formation of the pyramidal decussation (motor decussation), is called the lateral corticospinal (pyramidal) tract. Those fibers of the corticospinal tract that do not participate in the formation of the pyramidal decussation and do not pass to the opposite side continue their journey down as part of the anterior cord of the spinal cord. These fibers make up the anterior corticospinal (pyramidal) tract.
Then these fibers also pass to the opposite side, but through the white commissure of the spinal cord and end on the motor cells of the anterior horn of the opposite side of the spinal cord. It should be noted that all pyramidal paths are crossed, i.e. their fibers, on the way to the next neuron, sooner or later move to the opposite side. The second neuron of the descending voluntary motor pathway (corticospinal cord) are the cells of the anterior horns of the spinal cord, the long processes of which emerge from the spinal cord as part of the anterior roots and are sent as part of the spinal nerves to innervate skeletal muscles.
Extrapyramidal pathways , combined into one group, unlike the pyramidal pathways, have extensive connections in the brain stem and with the cerebral cortex, which has taken over the functions of monitoring and managing the extrapyramidal system. The cerebral cortex, which receives impulses both through direct (cortical direction) ascending sensory pathways and from subcortical centers, controls the motor functions of the body through extrapyramidal and pyramidal pathways.
The cerebral cortex influences the motor functions of the spinal cord through the cerebellum-red nuclei system, through the reticular formation, which has connections with the thalamus and striatum, and through the vestibular nuclei.
Thus, the centers of the extrapyramidal system include the red nuclei, one of the functions of which is to maintain muscle tone necessary to keep the body in a state of balance without effort of will. The red nucleus, which also belongs to the reticular formation, receives impulses from the cerebral cortex, the cerebellum (from the cerebellar proprioceptive pathways) and itself has connections with the motor nuclei of the anterior horns of the spinal cord. The red nucleus-spinal tract is part of the reflex arc, the afferent part of which is the spinal-cerebellar proprioceptive pathways. This pathway originates from the red nucleus (Monakov's bundle), passes to the opposite side ( Forel's decussation) and descends in the lateral funiculus of the spinal cord, ending on the motor cells of the spinal cord. The fibers of this pathway pass in the posterior part (tegmentum) of the pons and the lateral parts of the medulla oblongata.
An important link in the coordination of motor functions of the human body is the vestibulospinal tract . It connects the nuclei of the vestibular apparatus with the anterior horns of the spinal cord and provides adjustment reactions of the body in case of imbalance. of the vestibular nuclei take part in the formation of the vestibular-spinal tract . These fibers descend in the anterior funiculus of the spinal cord and end on the motor cells of the anterior horns of the spinal cord.
The nuclei that form the vestibulospinal tract are in direct connection with the cerebellum, as well as with the posterior longitudinal fasciculus, which in turn is connected with the nuclei of the oculomotor nerves. The presence of a connection with the nuclei of the oculomotor nerves ensures the preservation of the position of the eyeball (direction of the visual axis) when turning the head and neck.
In the formation of the posterior longitudinal fasciculus and those fibers that reach the anterior horns of the spinal cord (reticular-spinal tract), of the reticular formation take part , mainly the intermediate nucleus (nucleus of Cajal), the nucleus of the epithalamic (posterior) commissure (nucleus of Darkshevich ), to which fibers come from the basal ganglia of the cerebral hemispheres.
The functions of the cerebellum, which is involved in coordinating movements of the head, trunk and limbs and is in turn connected with the red nuclei and the vestibular apparatus, are controlled from the cerebral cortex through the bridge along the cortico-pontocerebellar pathway . This pathway consists of two neurons . The cell bodies of the first neuron lie in the cortex of the frontal, temporal, parietal and occipital lobes. Their processes, the cortico-pontine fibers, are directed to the internal capsule and pass through it. Fibers from the frontal lobe, which can be called frontopontine fibers, pass through the anterior limb of the internal capsule, nerve fibers from the temporal, parietal and occipital lobes pass through the posterior limb. Next, the fibers of the cortical-pontine tracts go through the base of the cerebral peduncle. In the anterior part (at the base) of the pons, the fibers of the corticopontine tracts end with synapses on the cells of the pons nuclei of the same side of the brain. The cells of the pontine nuclei with their processes constitute the second neuron of the corticocerebellar tract. The axons of the cells of the pons nuclei are folded into bundles - the transverse fibers of the pons, which pass to the opposite side, cross the descending bundles of fibers of the pyramidal tracts in the transverse direction and are directed through the middle cerebellar peduncle to the cerebellar hemisphere of the opposite side.
Thus, the pathways of the brain and spinal cord establish connections between afferent and efferent (effector) centers and participate in the formation of complex reflex arcs in the human body. Some pathways (fiber systems) begin or end in the brain stem and nuclei, providing functions that have a certain automaticity. These functions (for example, muscle tone, automatic reflex movements) are carried out without the participation of consciousness, although under the control of the cerebral cortex. Other pathways transmit impulses to the cerebral cortex, to the higher parts of the central nervous system, or from the cortex to the subcortical centers (to the basal ganglia, nuclei of the brain stem and spinal cord). The pathways functionally unite the body into one whole and ensure the consistency of its actions.
Test questions for the lecture:
1. General characteristics of motor pathways.
2. Structural and functional elements of the cortical-nuclear pathway.
3. Structural and functional elements of the cortical-nuclear pathway.
4. Structural and functional elements of the extrapyramidal system.
5. The role of subcortical structures in the formation of muscle tone.
6. The role of the structural elements of the midbrain in the regulation of muscle tone and the regulation of the quadrigeminal reflex.
Graphic basis for depicting conduction paths and reflex arcs
The diagram of pathways or reflex arcs requires a symbolic representation of the sections of the nervous system and the chain of neurons located in these sections.
Let's start with a picture of neurons. The first in the chain of neurons of the afferent pathway is the ganglion nerve cell, the body of which is always located in the peripheral part of the nervous system, in most cases in the sensitive nerve ganglion. In shape, a ganglion cell looks either like a bipolar neuron or (more often) like a pseudounipolar neuron (Fig. 1).
Rice. 1. Schematic representation of neurons and synaptic connections. Bipolar neuron (a); pseudounipolar neuron (b): 1 - receptor nerve ending, 2 - peripheral process (dendrite), 3 - neuron body, 4 - central process (axon), 5 - ending in contact with the second neuron in the chain, 6 - direction of impulse. Interneuron contact (c): 1 - first neuron, 2 - synapse, 3 - second neuron, 4 - direction of impulse. Motor neuron (d): 1 - neuron body, 2 - central process (axon), 3 - effector nerve ending, 4 - direction of impulse
The bodies of communication and motor neurons are located only in the central nervous system: communication - in centers of either the screen or nuclear type; motor - only in the motor nuclei (see Fig. 1).
For diagrams of most pathways and reflex arcs, a graphic representation of the parts of the nervous system is used (Fig. 2).