Роль коры больших полушарий
В задней центральной извилине, позади от центральной борозды, располагается зона кожной и суставно-мышечной чувствительности. Здесь воспринимаются и анализируются сигналы, возникающие при касании к нашему телу, при воздействии на него холода или тепла, болевых воздействиях.
В противоположность этой зоне — в передней центральной извилине, спереди от центральной борозды, расположена двигательная зона. В ней выявлены участки, которые обеспечивают движения нижних конечностей, мышц туловища, рук, головы. При раздражении этой зоны электротоком возникают сокращения соответствующих групп мышц. Ранения или другие повреждения коры двигательной зоны влекут за собой паралич мышц тела.
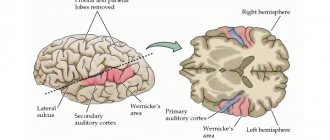
В височной доле находится слуховая зона. Сюда поступают и здесь анализируются импульсы, возникающие в рецепторах улитки внутреннего уха. Раздражения участков слуховой зоны вызывают ощущения звуков, а при поражении их болезнью утрачивается слух.
Зрительная зона расположена в коре затылочных долей полушарий. При ее раздражении электрическим током во время операций на мозге человек испытывает ощущения вспышек света и темноты. При поражении ее какой-либо болезнью ухудшается и теряется зрение.
Вблизи боковой борозды расположена вкусовая зона, где анализируются и формируются ощущения вкуса на основании сигналов, возникающих в рецепторах языка. Обонятельная зона расположена в так называемом обонятельном мозге, у основания полушарий. При раздражении этих зон во время хирургических операций или при воспалении люди ощущают запах или вкус каких-либо веществ.
Чисто речевой зоны не существует. Она представлена в коре височной доли, нижней лобной извилине слева, участках теменной доли. Их поражения болезнями сопровождаются расстройствами речи.
Большие полушария головного мозга занимают около 80% объема черепной коробки, и состоят из белого вещества, основа которого состоит из длинных миелиновых аксонов нейронов. Снаружи полушария покрывает серое вещество или кора головного мозга, состоящая из нейронов, безмиелиновых волокон и глиальных клеток, которые также содержатся в толще отделов этого органа.
Поверхность полушарий условно делится на несколько зон, функциональность которых заключается в управлении организмом на уровне рефлексов и инстинктов. Также в ней находятся центры высшей психической деятельности человека, обеспечивающие сознание, усвоение поступившей информации, позволяющей адаптироваться в окружающей среде, и через нее, на уровне подсознания, посредством гипоталамуса контролируется вегетативная нервная система (ВНС), управляющая органами кровообращения, дыхания, пищеварения, выделения, размножения, а также метаболизмом.
Для того чтобы разобраться что такое кора мозга и каким образом осуществляется ее работа, требуется изучить строение на клеточном уровне.
Функции
Кора занимает большую часть больших полушарий, а ее толщина не равномерна по всей поверхности. Такая особенность обусловлена большим количеством связующих каналов с центральной нервной системой (ЦНС), обеспечивающих функциональную организацию коры мозга.
Эта часть головного мозга начинает образовываться еще во время внутриутробного развития и совершенствуется на протяжении всей жизни, посредством получения и обработки сигналов, поступающих из окружающей среды. Таким образом, она отвечает за выполнение следующих функций головного мозга:
- связывает органы и системы организма между собой и окружающей средой, а также обеспечивает адекватную реакцию на изменения;
- обрабатывает поступившую информацию от моторных центров с помощью мыслительных и познавательных процессов;
- в ней формируется сознание, мышление, а также реализовывается интеллектуальный труд;
- осуществляет управление речевыми центрами и процессами, характеризующими психоэмоциональное состояние человека.
При этом данные поступают, обрабатываются, сохраняются благодаря значительному количеству импульсов, проходящих и образующихся в нейронах, связанных длинными отростками или аксонами. Уровень активности клеток можно определить по физиологическому и психическому состоянию организма и описать с помощью амплитудных и частотных показателей, так как природа этих сигналов похожа на электрические импульсы, а их плотность зависит от участка, в котором происходит психологический процесс.
До сих пор неясно, каким образом лобная часть коры больших полушарий влияет на работу организма, но известно, что она мало восприимчива к процессам, происходящим во внешней среде, поэтому все опыты с воздействием электрических импульсов на этот участок мозга, не находят яркого отклика в структурах. Однако отмечается, что люди, у которых лобная часть повреждена, испытывают проблемы в общении с другими индивидами, не могут реализовать себя в какой-либо трудовой деятельности, а также им безразличен их внешний вид и сторонние мнение. Иногда встречаются и другие нарушения в осуществлении функций этого органа:
- отсутствие концентрации внимания на предметах обихода;
- проявление творческой дисфункции;
- нарушения психоэмоционального состояния человека.
Поверхность коры полушарий поделена на 4 зоны, очерченные наиболее четкими и значимыми извилинами. Каждая из частей при этом контролирует основные функции коры головного мозга:
- теменная зона — отвечает за активную чувствительность и музыкальное восприятие;
- в затылочной части расположена первичная зрительная область;
- височная или темпоральная отвечает за речевые центры и восприятие звуков поступивших из внешней среды, кроме того участвует в формировании эмоциональных проявлений, таких как радость, злость, удовольствие и страх;
- лобная зона управляет двигательной и психической активностью, а также руководит речевой моторикой.
Быстро менять свое поведение нам помогает премоторная кора
Долгосрочное обучение – такое, как изучение языка, математики или новых движений, вероятно, требует кортикальной пластичности: изменений в структуре мозга, но часто нам требуются гораздо более быстрые изменения, иногда даже после единичных ошибок. Группа профессора Ли Миллера из лаборатории контроля движения конечностей Северо-Западного Университета в статье, опубликованной в одном из топовых нейрожурналов, Neuron, предложила нейронный механизм для быстрого развития нового моторного навыка без изменения функциональной связи внутри или между кортикальными областями. Ли Миллер будет приглашенным спикером конференции BCISamara-2019, которая состоится в октябре и наш портал обязательно побеседует с ученым о новых разработках, а пока давайте расскажем о его статье.
Графический абстракт работы. Miller et al, Neuron, 2018
Как пишут авторы, один из самых фундаментальных вопросов в нейробиологии заключается в том, как скоординированная деятельность взаимосвязанных нейронов порождает поведение и как эти нейроны быстро и гибко изменяют свой «выходящий сигнал» во время обучения для адаптации поведения к изменившимся условиям. Имеются данные о том, что обучение, продолжающееся от нескольких дней до нескольких недель, связано с постоянными синаптическими изменениями в коре головного мозга (Kleim et al., 2004). Тем не менее, поведение также может быть адаптировано гораздо быстрее: двигательные ошибки могут быть исправлены на основе последовательных проб (trial-a-trial basis, Thoroughman and Shadmehr, 2000), а сенсорные ассоциации могут быть выучены даже после одного воздействия (Bailey and Chen, 1988).
Есть еще один факт: существуют ограничения на типы быстрого моторного обучения. В эксперименте с интерфейсом мозг-компьютер обезьяны с трудом научились управлять курсором компьютера, когда новый декодер управления потребовал, чтобы они изменили естественную сопряженную изменчивость между нейронами, активность которых считывали электроды (Sadtler et al, 2014). Судя по всему, такие ковариационные (то есть, одновременно изменяющие активность) структуры связана с синаптической связностью (простите за тавтологию), которая не так-то легко меняется на временных масштабах от секунд до минут (Okun et al., 2015). Таким образом все эти данные говорят о том, что когда мы очень быстро меняем свое поведение, изменения в коннектоме не могут быть основным механизмом этого.
Что же сделали авторы работы? В их эксперименте две черные макаки выполняли «стандартный» тест «от центра к краю»: на экране курсор появлялся в центральной точке, а затем обезьяна должна была при помощи манипулятора привести его к одной из равномерно распределенных по кругу восьми мишеней на краю, которые появлялись после звукового сигнала. На достижение мишени давалась секунда, и еще полсекунды курсор нужно было удержать. Однако после первого теста к манипулятору особые сервоприводы прикладывали крутящие усилия, и животному требовалось скорректировать движение, чтобы выполнить условие задачи. И обезьянки достаточно быстро справлялись с задачей.
Траектории движения курсора во время разных задач
Одновременно авторы записывали активность двух участков мозга – дорсальной премоторной коры (PMd), в которой происходит планирование движения и основной моторной коры (M1), которая, собственно, и осуществляет движение.
Исследователи выдвинули четыре гипотезы – где же происходит обучение (см. рисунок ниже), исходя из пути сигнала: от зрительной коры к PMd, затем в M1 и затем к собственно движению. Первая гипотеза предполагала научение еще до премоторной коры, затем на уровне «нулевого планирования» в премоторной коре, затем – на уровне карты взаимодействия PMd-M1 и на уровне локальной связности. Каждой гипотезе соответствовала своя расчетная модель.
Четыре гипотезы, выдвинутые авторами и связанные с ними модели
Анализ данных, полученных во время краткосрочной адаптации к двум разным тестам, которая достигалась за один день, показал очень интересные результаты: авторы не увидели никаких изменений как в коннективности внутри каждой из областей, так и в нейронной связаной изменчивости (ковариантности) между зонами PMd и M1. Оказалось, что адаптация к изменяющимся условиям происходила в особом подпространстве «output-NULL» премоторной коры, которое постепенно регулировало выход в нижележащую область цепочки с полным сохранением связности.
Текст: Алексей Паевский
Perich, M. G., Gallego, J. A., & Miller, L. E. (2018). A Neural Population Mechanism for Rapid Learning. Neuron.doi:10.1016/j.neuron.2018.09.030
Литература:
Kleim, J.A., Hogg, T.M., VandenBerg, P.M., Cooper, N.R., Bruneau, R., and Remple, M. (2004). Cortical synaptogenesis and motor map reorganization occur during late, but not early, phase of motor skill learning. J. Neurosci. 24, 628–633.
Thoroughman, K.A., and Shadmehr, R. (2000). Learning of action through adaptive combination of motor primitives. Nature 407, 742–747.
Bailey, C.H., and Chen, M. (1988). Morphological basis of short-term habituation in Aplysia. J. Neurosci. 8, 2452–2459.
Sadtler, P.T., Quick, K.M., Golub, M.D., Chase, S.M., Ryu, S.I., Tyler-Kabara, E.C., Yu, B.M., and Batista, A.P. (2014). Neural constraints on learning. Nature 512, 423–426.
Okun, M., Steinmetz, N., Cossell, L., Iacaruso, M.F., Ko, H., Bartho´ , P., Moore, T., Hofer, S.B., Mrsic-Flogel, T.D., Carandini, M., and Harris, K.D. (2015). Diverse coupling of neurons to populations in sensory cortex. Nature 521, 511–515.
Первая и вторая сигнальные системы
Неоценима роль коры большого мозга в совершенствовании первой сигнальной системы и развитии второй. Эти понятия разработаны И.П.Павловым. Под сигнальной системой в целом понимают всю совокупность процессов нервной системы, осуществляющих восприятие, переработку информации и ответную реакцию организма. Она связывает организм с внешним миром.
Первая сигнальная система обусловливает восприятие посредством органов чувств чувственно-конкретных образов. Она является основой для образования условных рефлексов. Эта система существует как у животных, так и у человека.
В высшей нервной деятельности человека развилась надстройка в виде второй сигнальной системы. Она свойственна только человеку и проявляется словесным общением, речью, понятиями. С появлением этой сигнальной системы стали возможными отвлеченное мышление, обобщение бесчисленных сигналов первой сигнальной системы. По И.П.Павлову, слова превратились в «сигналы сигналов».
Возникновение второй сигнальной системы стало возможным благодаря сложным трудовым взаимоотношениям между людьми, так как эта система является средством общения, коллективного труда. Словесное общение не развивается вне общества. Вторая сигнальная система породила отвлеченное (абстрактное) мышление, письмо, чтение, счет.
Слова воспринимаются и животными, но совершенно отлично от людей. Они воспринимают их как звуки, а не их смысловое значение, как люди. Следовательно, у животных нет второй сигнальной системы. Обе сигнальные системы человека взаимосвязаны. Они организуют поведение человека в широком смысле слова. Причем вторая изменила первую сигнальную систему, так как реакции первой стали в значительной мере зависеть от социальной среды. Человек стал в состоянии управлять своими безусловными рефлексами, инстинктами, т.е. первой сигнальной системой.
Функции коры мозга
Знакомство с наиболее важными физиологическими функциями коры большого мозга свидетельствует о необычайном ее значении в жизнедеятельности. Кора вместе с ближайшими к ней подкорковыми образованиями является отделом центральной нервной системы животных и человека.
Функции коры головного мозга — осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения) человека. Не случайно у него она получила наибольшее развитие. Исключительным свойством коры являются сознание (мышление, память), вторая сигнальная система (речь), высокая организация труда и жизни в целом.
Особенности строения коры мозга
Анатомическое строение коры больших полушарий обусловливает ее особенности и позволяет выполнять возложенные на нее функции. Кора головного мозга владеет следующим рядом отличительных черт:
- нейроны в ее толще располагаются послойно;
- нервные центры находятся в конкретном месте и отвечают за деятельность определенного участка организма;
- уровень активности коры зависит от влияния ее подкорковых структур;
- она имеет связи со всеми нижележащими структурами центральной нервной системы;
- наличие полей разных по клеточному строению, что подтверждается гистологическим исследованием, при этом каждое поле отвечает за выполнение какой-либо высшей нервно деятельности;
- присутствие специализированных ассоциативных областей позволяет устанавливать причинно-следственную связь между внешними раздражителями и ответом организма на них;
- способность к замещению поврежденных участков близлежащими структурами;
- этот отдел мозга способен сохранять следы возбуждения нейронов.
Большие полушария головного мозга состоят главным образом из длинных аксонов, а также содержит в своей толще скопления нейронов, образующих наибольшие ядра основания, которые входят в состав экстрапирамидальной системы.
Как уже говорилось, формирование коры мозга происходит еще во время внутриутробного развития, причем вначале кора состоит из нижнего слоя клеток, а уже в 6 месяцев ребенка в ней сформированы все структуры и поля. Окончательное становление нейронов происходит к 7-летнему возрасту, а рост их тел завершается в 18 лет.
Интересен тот факт, что толщина коры не равномерна на всей протяженности и включает в себя разное количество слоев: например, в области центральной извилины она достигает своего максимального размера и насчитывает все 6 слоев, а участки старой и древней коры имеют 2-х и 3-х слойное строение соответственно.
Нейроны этой части мозга запрограммированы на восстановление поврежденного участка посредством синоптических контактов, таким образом каждая из клеток активно старается восстановить поврежденные связи, что обеспечивает пластичность нейронных корковых сетей. Например, при удалении или дисфункции мозжечка, нейроны, связывающие его с конечным отделом, начинают прорастать в кору больших полушарий.
анализатора, замыкательного аппарата условно-рефлексных связей и рабочего прибора. Слабость замыкательной функции коры и следовых проявлений можно наблюдать у детей с выраженной умственной отсталостью, когда образовавшиеся условные связи между нейронами хрупки и ненадежны, что влечет за собой трудности в обучении.
Кора головного мозга включает в себя 11 областей, состоящих из 53 полей, каждому из которых в нейрофизиологии присвоен свой номер.
Области и зоны коры
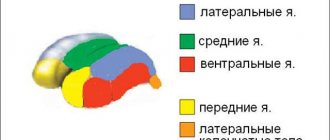
Кора относительно молодая часть ЦНС, развывшаяся из конечного отдела мозга. Эволюционно становление этого органа происходило поэтапно, поэтому ее принято разделять на 4 типа:
- Архикортекс или древняя кора в связи с атрофией обоняния превратился в гиппокамповую формацию и состоит из гиппокампа и сопряженных ему структур. С помощью ее регулируется поведение, чувства и память.
- Палеокортекс или старая кора, составляет основную часть обонятельной зоны.
- Неокортекс или новая кора имеет толщину слоя около 3—4 мм. Является функциональной частью и совершает высшую нервную деятельность: обрабатывает сенсорную информацию, отдает моторные команды, а также в ней формируется осознанное мышление и речь человека.
- Мезокортекс является промежуточным вариантом первых 3 типов коры.
Кора головного мозга имеет сложную анатомическую структуру и включает в себя сенсорные клетки, моторные нейроны и интернероны, обладающих способностью останавливать сигнал и возбуждаться в зависимости от поступивших данных. Организация этой части мозга построена по колончатому принципу, в котором колонки делаться на микромодули, имеющие однородное строение.
Основу системы микромодулей составляют звездчатые клетки и их аксоны, при этом все нейроны одинаково реагируют на поступивший афферентный импульс и посылают также синхронно в ответ эфферентный сигнал.
Формирование условных рефлексов, обеспечивающих полноценное функционирование организма, и происходит благодаря связи головного мозга с нейронами, расположенными в различных частях тела, а кора обеспечивает синхронизацию умственной деятельности с моторикой органов и областью, отвечающей за анализ поступающих сигналов.
Передача сигнала в горизонтальном направлении происходит через поперечные волокна, находящиеся в толще коры, и передают импульс от одной колонки к другой. По принципу горизонтальной ориентации кору мозга можно поделить на следующие области:
- ассоциативная;
- сенсорная (чувствительная);
- моторная.
При изучении этих зон применялись различные способы воздействия на нейроны, входящие в ее состав: химическое и физическое раздражение, частичное удаление участков, а также выработка условных рефлексов и регистрация биотоков.
Ассоциативная зона связывает поступившую сенсорную информацию с полученными ранее знаниями. После обработки формирует сигнал и передает его в двигательную зону. Таким образом она участвует в запоминании, мышлении и обучении новым навыкам. Ассоциативные участки коры головного мозга расположены в близости с соответствующей сенсорной зоной.
Чувствительная или сенсорная зона занимает 20% коры головного мозга. Она также состоит из нескольких составляющих:
- соматосенсорной, расположенной в теменной зоне отвечает за тактильную и вегетативную чувствительность;
- зрительной;
- слуховой;
- вкусовой;
- обонятельной.
Импульсы от конечностей и органов осязания левой стороны тела, поступают по афферентным путям в противоположную долю больших полушарий для последующей обработки.
Нейроны моторной зоны возбуждаются при помощи импульсов, поступивших от клеток мускулатуры, и находятся в центральной извилине лобной доли. Механизм поступления данных схож с механизмом сенсорной зоны, так как двигательные пути образуют перехлест в продолговатом мозге и следуют в расположенную напротив моторную зону.
Кора больших полушарий образована несколькими слоями нейронов. Характерной особенностью этой части мозга является большое количество морщин или извилин, благодаря чему ее площадь во много раз превосходит площадь поверхности полушарий.
Корковые архитектонические поля определяют функциональное строение участков коры головного мозга. Все они различны по морфологическим признакам и регулируют разные функции. Таким образом выделяется 52 различных поля, расположенных на определенных участках. По Бродману это разделение выглядит следующим образом:
- Центральная борозда разделяет лобную долю от теменной области, впереди нее пролегает предцентральная извилина, а сзади — позадицентральная.
- Боковая борозда отгораживает теменную зону от затылочной. Если развести ее боковые края то внутри можно рассмотреть ямку, в центре которой имеется островок.
- Теменно-затылочная борозда отделяет теменную долю от затылочной.
В предцентральной извилине расположено ядро двигательного анализатора, при этом к мышцам нижней конечности относятся верхние части передней центральной извилины, а к мышцам полости рта, глотки и гортани – нижние.
Правосторонняя извилина образует связь с двигательным аппаратом левой половины тела, левосторонняя – с правой частью.
В позадицентральной извилине 1 доли полушария содержится ядро анализатора тактильных ощущений и она также связана с противолежащей частью тела.
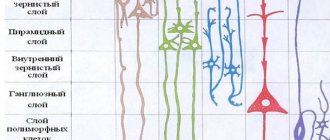
Клеточные слои
Кора головного мозга осуществляет свои функции посредством нейронов, находящихся в ее толще. Причем количество слоев этих клеток может отличаться в зависимости от участка, габариты которых также разнятся по размеру и топографии. Специалисты выделяют следующие слои коры головного мозга:
- Поверхностный молекулярный сформирован в основном из дендритов, с небольшим вкраплением нейронов, отростки которых не покидают границы слоя.
- Наружный зернистый состоит из пирамидальных и звездчатых нейронов, отростки которых связывают его со следующим слоем.
- Пирамидальный образован пирамидными нейронами, аксоны которых направлены вниз, где обрываются или образуют ассоциативные волокна, а дендриты их соединяют этот слой с предыдущим.
- Внутренний зернистый слой сформирован звездчатыми и малыми пирамидальными нейронами, дендриты которых уходят в пирамидальный слой, а также его длинные волокна уходят в верхние слои или спускаются вниз в белое вещество мозга.
- Ганглионарный состоит из крупных пирамидальных нейроцитов, их аксоны выходят за пределы коры и связывают различные структуры и отделы ЦНС между собой.
Мультиформный слой сформирован всеми видами нейронов, а их дендриты ориентированы в молекулярный слой, а аксоны пронизывают предыдущие слои или выходят за пределы коры и образуют ассоциативные волокна, образующие связь клеток серого вещества с остальными функциональными центрами головного мозга.
ЗОНЫ БОЛЬШИХ ПОЛУШАРИЙ
Локализация функций в больших полушариях. Кора больших полушарий головного мозга делится на основные зоны, состоящие из нескольких корковых полей. Каждая из этих зон выполняет определенную общую функцию, а составляющие ее поля специализированно участвуют в реализации отдельных элементов этой функции. Однако благодаря проводящим путям в осуществлении отдельных звеньев высшей и низшей нервной деятельности участвует несколько зон больших полушарий, определенные подкорковые центры, ядра мозгового ствола и сегменты спинного мозга.
При тонкой и точной специализации определенных групп нейронов головной и спинной мозг функционируют как единое целое. Психические функции головного мозга также не ограничены отдельными участками коры, а являются результатом совместной деятельности обширных зон больших полушарий и подкорковых центров.
Рис. 123. Индивидуальные изменения основных полей новой коры больших полушарий у трех взрослых (А, Б, В). Цифры— поля по Бродману Моторная зона (поле 4) расположена в передней центральной извилине вдоль центральной борозды. В верхней четверти зоны находятся двигательные центры для мышц ног.
Сверху расположены нейроны, иннервирующие мышцы пальцев ног, а снизу — бедра и туловища. Две средние четверти заняты центрами для рук, выше — центр мышц лопатки, а ниже — мышц пальцев. И, наконец, в нижней четверти передней центральной извилины находятся центры мышц лица и речевого аппарата. В результате исторического развития головного мозга человека в процессе труда и речи особенно большое место занимают группы нейронов, которые вызывают сокращение мышц кисти руки, главным образом большого пальца, и мышц лица, языка и гортани. К ним поступают центростремительные волокна из проприорецепторов, входящие по задним корешкам в спинной мозг, где они поднимаются в составе заднего столба той же стороны до ядер нежного и клиновидного пучков продолговатого мозга. Из этих ядер выходят волокна вторых нейронов, образующие медиальную петлю и после перекреста достигающие ядер зрительного бугра противоположной стороны. Отсюда большая часть центростремительных волокон третьих нейронов достигает задней центральной извилины и далее поступает в переднюю центральную извилину, а меньшая часть входит в нее прямым путем. Таким образом, передняя центральная извилина посредством волокон, проходящих в проводящих путях коры, связана с задней центральной извилиной. Из моторной зоны выходят центробежные двигательные волокна пирамидных нейронов, которые составляют пирамидные проводящие пути; они достигают нейронов передних рогов спинного мозга. Моторная зона вызывает координированные движения скелетных мыщц, преимущественно на противоположной стороне тела. Она функционирует совместно с подкорковыми центрами — полосатыми телами, а также люисовым телом, красным ядром и черным веществом. При поражениях определенных участков передней центральной извилины нарушаются произвольные движения отдельных групп мышц. Неполное поражение зоны вызывает нарушение движений— парез, а полное ее разрушение — паралич. Зона кожно-мышечной чувствительности (поля 1, 2, 3, 43 и частично 5 и 7) расположена в задней центральной извилине вдоль задней центральной борозды. В этой зоне особенно сильно развиты зернистые слои коры, к которым подходят центростремительные волокна из рецепторов кожи, идущие в составе тех же проводящих путей, как и волокна из проприорецепторов. Расположение воспринимающих групп нейронов такое же, как в моторной зоне. Наибольшую поверхность занимают нейроны, воспринимающие импульсы из рецепторов кисти руки, лица, языка и гортани. Поле 7 больше других полей связано с чувствительностью руки. Зона кожно-мышечной чувствительности не полностью отграничена от моторной зоны, так как в полях 3, 4 и 5 происходит сочетание зернистых нейронов с гигантскими пирамидными нейронами. В моторной зоне находится примерно 80% двигательных нейронов, а в зоне кожно-мышечной чувствительности — 20%. В каждое полушарие поступают импульсы главным образом из рецепторов противоположной стороны тела, но также и из рецепторов той же стороны. В эту зону поступают центростремительные импульсы преимущественно из бокового и полулунного ядер зрительного бугра. При поражениях определенных участков задней центральной извилины нарушается чувствительность в отдельных участках кожи. Потеря способности узнавать предметы при их осязании обозначается как тактильная агнозия. При нарушениях функций зоны наблюдаются расстройства осязания, болевых и температурных ощущений кожи и мышечно-суставной чувствительности. Неполное поражение зоны вызывает понижение рецепции — гипостезию, а полное — ее потерю — анестезию. Лобная зона (поля 6, 5, 9, 10, 11, 44, 45, 46, 47) расположена в лобной доле впереди моторной. Она делится на премоторную и речедвигательную. Премоторная зона (поля 6, 8, 9, 10, 11) регулирует тонус скелетных мышц и координированные движения тела, ориентирующие его в пространстве. С полем 10, которое участвует в выполнении двигательных условных рефлексов, функционально связано поле 46. В премоторную зону поступают центростремительные импульсы из внутренних органов и из нее исходит значительная часть центробежных вегетативных волокон. Поэтому поражение премоторной зоны вызывает нарушение координации движений — атаксию и расстройства функций сердечнососудистой, дыхательной, пищеварительной и других систем внутренних органов. Зрительная зона (поля 17, 18, 19) расположена на внутренней поверхности затылочной доли по обеим сторонам шпорной борозды. У человека она занимает 12% общей поверхности коры. Поле 17 находится на затылочном полюсе; оно окружено полем 18, которое окружает поле 19, граничащее с задним отделом лимбической области, верхней и нижней теменными областями. В поле 17 — центральном поле зрительной зоны в 16 раз больше нейронов, чем в центральном поле слуховой зоны (поле 41), и в 10 раз больше нейронов, чем в центральном поле моторной зоны (поле 4). Это указывает на ведущее в историческом и индивидуальном развитии человека значение зрения. Из сетчатки 900 тыс.— 1 млн. центростремительных волокон зрительных нервов доходит до наружного коленчатого тела, в котором точно проецируются отдельные части сетчатки. Центростремительные волокна нейронов наружного коленчатого тела направляются в зрительную зону, преимущественно в основное зрительное поле 17. Другими промежуточными зрительными центрами, участвующими в передаче не зрительных импульсов, а глазодвигательных, являются подушка зрительного бугра и передние бугры четверохолмия. До поступления в наружное коленчатое тело волокна зрительного нерва перекрещиваются. Благодаря этому перекресту в составе зрительного пути, направляющегося в зрительную зону каждого полушария, 50% волокон своей стороны и 50% волокон противоположной стороны. В зрительную зону левого полушария поступают зрительные импульсы из левых половин сетчаток обоих глаз, а в зону правого полушария — из правых половин сетчаток обоих глаз. Поэтому разрушение одной из зрительных зон вызывает слепоту в одноименных половинах сетчаток в обоих глазах — гемианопсию. В зрительных нервах, кроме центростремительных волокон, проходят и несколько более толстые центробежные волокна к мышцам радужной оболочки и центробежные тонкие симпатические волокна из нейронов подкорковых центров. Небольшая часть центростремительных волокон зрительного нерва не прерывается в подкорковых образованиях, а прямо направляется в мозжечок и зрительные зоны больших полушарий. Разрушение обоих полей 17 вызывает полную корковую слепоту, разрушение поля 18 приводит к потере зрительной памяти при сохранении зрения, что обозначается как зрительная агнозия, а разрушение поля 19 — к потере ориентации в непривычной обстановке. Слуховая зона (поля 41, 42, 21, 22, 20, 37) расположена на поверхности височной доли, преимущественно передней поперечной височной извилины и верхней височной извилины. Поле 41, расположенное в верхней височной извилине и в передней части поперечной извилины, является проекцией кортиева органа улитки. Из органа Корти центростремительные импульсы проходят через спиральный узел по улиточному нерву, состоящему примерно из 30 тыс. волокон. В этом узле находятся первые биполярные нейроны слухового пути. Далее волокна первых нейронов передают слуховые импульсы в ядра слухового нерва в продолговатом мозге, где находятся вторые нейроны. Волокна ядер слухового нерва связываются с ядрами лицевого нерва в продолговатом мозге и глазодвигательного нерва в передних буграх среднего мозга. Поэтому при сильных звуках рефлекторно сокращаются мышцы лица, век, ушной раковины и вызываются движения глаз. Большая часть волокон ядер слухового нерва перекрещивается в варолиевом мосту, а меньшая проходит на своей стороне. Затем волокна слухового пути поступают в боковую лемнисковую петлю, которая заканчивается в задних буграх четверохолмия и во внутреннем коленчатом теле, где находятся третьи нейроны — их волокна проводят центростремительные импульсы в слуховую зону. Существуют также прямые пути, связывающие ядра слуховых нервов с мозжечком и слуховой зоной. Большая часть прямых мозжечковых путей образуется вестибулярным нервом, а меньшая— улитковым нервом, составляющими вместе общий ствол слухового нерва. Вестибулярный аппарат проецируется также в слуховой зоне. Разрушение поля 41 на одной стороне вызывает глухоту на противоположной стороне и ослабление слуха на своей стороне, а разрушение полей 41 на обеих сторонах ведет к полной корковой глухоте. Разрушение поля 22 в передней трети верхней височной извилины приводит к музыкальной глухоте — теряется восприятие интенсивности тона, тембра и ритма звуков — слуховая агнозия. Разрушение полей 21 и 20 в средней и нижней височных извилинах вызывает атаксию — расстройство равновесия и координации движений. В слуховой зоне расположен также рече-слуховой центр. Обонятельная и вкусовая зоны. Обонятельная зона находится в древней коре, в которую поступают центростремительные импульсы из обонятельных клеток. Кроме обонятельной функции, она выполняет также вкусовую и участвует в деятельности пищеварительной, выделительной и половой систем. Раньше считали, что гиппокамп выполняет обонятельную функцию. В настоящее время полагают, что вместе с лимбической системой, гипоталамической областью промежуточного мозга и гипофизом, средним и продолговатым мозгом и особенно ретикулярной формацией гиппокамп участвует в общих двигательных реакциях и вегетативных рефлексах при эмоциях. Собственно вкусовая зона, вероятно, расположена в поле 43, которое находится в нижнем отделе задней центральной извилины.
Лимбическая извилина (заднее поле 23 и переднее поле 24) и кора островка (поля 13 и 14) участвуют в высшей нервной деятельности. Все зоны коры не обособлены, а связаны между собой проводящими путями. Центры речи (поля 44, 45, 46, 39, 40, 42, 22,37). Двигательный центр речи расположен в нижней части передней центральной извилины в поле 44. У большинства правшей площадь поля 44 в левом полушарии больше, чем в правом полушарии. Поле 44 вызывает сложные сокращения речевой мускулатуры, необходимые для произнесения слов. При разрушении этого поля человек не может говорить, но может производить простейшие сокращения речевой мускулатуры — кричать и петь. Это моторная, двигательная афазия, которая в некоторых случаях проявляется в отсутствии сокращений мышц языка и остальной речевой мускулатуры. Так как в этих случаях слуховой центр речи не поврежден, то понимание речи окружающих сохраняется. При поражении поля 44 часто нарушается не только устная речь, но и внутренняя речь или способность формулировать мысли словами без их произнесения, на основе накопленных звуковых образов, имеющих определенное смысловое содержание. При этом затруднено чтение про себя, расстроена способность писать произвольно и под диктовку, но сохранено копирование букв при письме. У правшей моторная афазия наблюдается при поражении левого полушария, а левшей — правого.
Рис. 129. Локализация центров речи: 1 — двигательный, 2 — слуховой, 3 — зрительный Впереди поля 44 расположено поле 45, которое регулирует построение грамматически правильных сочетаний слов и пение. При поражении этого поля вследствие потери памяти на приемы произношения пение расстраивается. Мимика и жестикуляция, придающие речи ее выразительность, осуществляются благодаря импульсам, поступающим из поля 46 в поля 44 и 45, в поля премоторной области и в подкорковые центры. Слуховой, или сенсорный, центр речи расположен в заднем отделе левой верхней височной извилины в поле 42, которое осуществляет понимание слова при слышании его. Если поле разрушается, теряется способность понимания смысла слов, но сохраняется их восприятие как звуков — сенсорная афазия, или речевая глухота. При этом вследствие отсутствия понимания собственной речи, иногда наблюдается чрезмерная говорливость — логоррея, или словесный понос. В задней части поля 22 фиксируются связи звуковых образов слов со всеми воспринимающими зонами, в которых возникают представления о предметах и явлениях. Поэтому поражение этого поля также вызывает сенсорную афазию. Поля 39 и 40, расположенные в теменной доле рядом с полем 22, осуществляют понимание смысла сочетаний слов или фраз. Поэтому их поражение приводит к расстройству речи, которое называется семантической (смысловой) афазией. При поражении поля 39, вследствие потери способности узнавать буквы и цифры и понимать смысл видимых письменных образов слов и цифр, теряется способность читать вслух, писать и считать. Поражение поля 40 вызывает потерю способности писать, так как отсутствует ориентация движений в пространстве и нарушена их последовательность. Это отсутствие способности производить системные, целенаправленные движения (апраксия) не исключает возможности правильно совершать отдельные движения руки, не связанные с письмом. Следовательно, процесс письма у правшей осуществляется, височной, нижнетеменной и нижнелобной областями левого полушария. При поражении поля 37 вызывается потеря памяти на слова — амнестическая афазия. Таким образом, в осуществлении функции речи участвуют большие полушария головного мозга в целом, но особенная роль выполняется отдельными полями коры. У правшей в результате преимущественного развития функций правой руки и правой половины тела особенно развиты сложнейшие психические функции левого полушария головного мозга.
Похожие материалы:
Филогенез головного мозга
Строение полушарий головного мозга
Кровоснабжение головного мозга
Спинномозговая жидкость