In the medulla oblongata, the axons of second neurons pass between the inferior olive and the inferior cerebellar peduncle. In the tegmentum of the bridge they approach its dorsal surface. At the border with the midbrain, the fibers of the anterior spinocerebellar tract turn dorsally into the region of the superior medullary velum. Here, part of the fibers once again passes to the opposite side, and then through the superior cerebellar peduncles (pedunculi cerebellares superiores) they reach the anterosuperior parts of the cerebellum and enter the nucleus of the tent, nucleus fastigii, cerebellar vermis, vermis cerebelli. ). The tent nucleus is a collector of afferent information. From here it goes to the piriform neurons of the cerebellar cortex - Purkinje cells. Jan Evangelista Purkinje, 1787-1869, Czech physiologist, histologist, embryologist and pharmacologist. Pear-shaped neurons process the received information and transmit it to the dentate nucleus (nucleus dentatus) of the cerebellar hemisphere of the same name. The afferent pathways end in this structure. The dentate nucleus is the final link of the cerebellar regulators. Its neurons form and send efferent (control) signals through the superior cerebellar peduncles to the red nucleus of the midbrain on the opposite side (Wernecking's chiasm). It is possible to trace the fiber systems through which information from the vermis cortex reaches the red nucleus, the cerebellar hemisphere, as well as the overlying parts of the brain - the cerebral cortex. From the cortex of the vermis, through the corky and globular nuclei, information through the superior cerebellar peduncle is sent to the red nucleus of the opposite side (cerebellotegmental tract). The vermis cortex is connected by associative fibers to the cerebellar hemisphere cortex, from where information enters the dentate nucleus of the cerebellum. There are also connections between the cerebellum and the cortex through the thalamus. In particular, the axons of the neurons of the dentate nucleus exit through the superior cerebellar peduncle into the tegmentum of the bridge, pass to the opposite side and go to the thalamus. Having switched to the next neuron in the thalamus, the paths follow to the cerebral cortex, to the postcentral gyrus. The spinocerebellar tracts described are the main ones, but not the only known spinocerebellar tracts. The sphenocerebellar tract (tractus cuneocerebellaris) is considered part of the posterior spinocerebellar tract. The rostral spinocerebellar tract (tractus spinocerebellaris rostralis) is considered part of the anterior spinocerebellar tract. There are also indirect spinocerebellar tracts. Among them, the most studied are: spino-olivary-cerebellar tracts, spino-bulbo-cerebellar tracts and spino-reticulo-cerebellar tracts.
Thanks to information arriving along the anterior spinal cerebellar tracts and posterior spinal cerebellar tracts, the cerebellum, as one of the nerve centers in the hierarchy of regulators of the musculoskeletal system, is involved in controlling movements that ensure body balance without the participation of higher parts of the brain (cerebral cortex) brain), without the participation of consciousness.
See: Neurology: Dictionary, Neurology: Internet Resources.
| LIBRARY = LIBRARY Human Physiology = Human Physiology, Human Anatomy = Human Anatomy, Human Biochemistry = Human Biochemistry, Human Psychology = Human Psychology, Medicine = Medicine, Mathematics = Mathematics, Chemistry = Chemistry, Physics = Physics, General Scientific Literature = General Science Lexis. Click here to access any source in the site's library! Click here and receive access to the any reference of the library! “I AM LEARNED. . . N E D O U C H A ?” T E S T V A S H E G O I N T E L L E C T A Premise : The effectiveness of the development of any branch of knowledge is determined by the degree of compliance with the methodology of knowledge - the knowable entity. |
Error? Click here and fix it! Search on the site E-mail of the author (author)
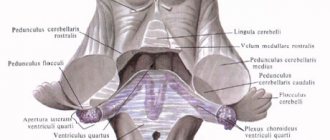
Pathways of proprioceptive sense of the cerebellar direction (Flexig and Govers)
⇐ PreviousPage 2 of 36Next ⇒
It has long been believed that the cerebellum is one of the centers of coordination and synergy of movements, regulation of muscle tone, and maintaining balance. Academician L.A. Orbeli came to the conclusion that “the cerebellum is the highest adaptation-trophic center,” where the integration of somato-vegetative functions occurs.
Currently, the cerebellum is considered as a unique system that coordinates motor acts. Thanks to its extensive connections, the cerebellum receives information from receptors located in muscles, tendons, joints, as well as from the vestibular, visual and auditory analyzers, and from the reticular formation. This enormous influx of sensory information ultimately serves to regulate motor functions. Integrated information is issued in the form of efferent signals to the leading motor centers of the spinal cord and brain.
Apparently, the cerebellum does not play a leading role in organizing motor acts, does not program movements, but optimizes movement programs as they are implemented. The cerebellum takes part in the control of movements, making appropriate adjustments to the activity of the motor centers of the brain and spinal cord. It is assumed that the cerebellum not only regulates the activity of individual motor centers, but also coordinates their work.
Consequently, the most important afferent connections of the cerebellum are proprioceptive pathways from the spinal cord, vestibular nuclei, extrapyramidal system, motor centers of the cerebral cortex; the most important efferent connections are the paths to the motor centers of the brain and spinal cord.
The cerebellum receives information about the state and activity of the muscular-articular system through several channels. These are, firstly, direct spinocerebellar tracts, directly connecting the spinal cord with the cerebellum, and, secondly, conductors that switch on the way to the cerebellum in the nuclei of the brain stem. These nuclei are called “precerebellar”.
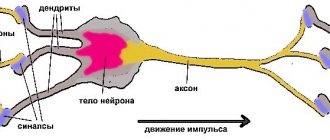
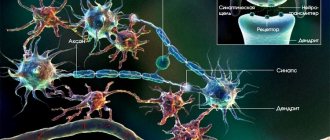
Proprioceptors are located in muscles, tendons, articular surfaces, joint capsules, ligaments, fascia, and periosteum. Irritations are transmitted along the sensory fibers of the spinal nerves to the cells of the spinal ganglia (I neuron) and from them along the fibers of the dorsal roots into the gray matter of the spinal cord. This is where two anatomically separate nerve pathways originate:
A. Posterior spinocerebellar tract (tractus spinocerebellaris posterior)
(Flexig's bundle) starts from the cells of the thoracic nucleus
(nucleus thoracicus)
(II neuron). The fibers of this path pass, without forming a decussation, along the periphery of the lateral cord of the spinal cord, continue into the medulla oblongata and enter the lower cerebellar peduncles. The path ends in the cortex of the cerebellar vermis on its side (paleocerebellum).
B. Anterior spinocerebellar tract (tractus spinocerebellaris anterior)
(Gowers bundle) originates from the cells of the intermediate-medial nucleus
(substantia intermedia medialis)
(II neuron). The fibers of the pathway predominantly pass to the opposite side in the anterior white commissure of the spinal cord (first decussation) and along the periphery of the lateral funiculus go in an ascending direction, pass through the medulla oblongata and the pons and reach the midbrain. The anterior spinocerebellar tract then passes along the superior cerebellar peduncle, forming a second (partial) decussation in the anterior medullary velum, and similarly to the posterior tract ends in the cortex of the cerebellar vermis.
Structural differences between the two spinocerebellar tracts determine their functional differences. It is believed that through the posterior pathway the cerebellum receives information from the muscle and tendon receptors of each separate muscle group. The anterior pathway into the cerebellum receives signals, apparently from large muscle groups.
The second type of connections “musculoskeletal system - spinal cord - cerebellum”, switching on the nuclei of the brain stem, includes the posterior external arcuate fibers.
As part of the lower legs, reticular-cerebellar fibers pass to the cortex of the cerebellar hemispheres, which close the spinal-reticular-cerebellar connections. It is assumed that through them the cerebellum receives additional proprioceptive information about the movements performed by the skeletal muscles of the limbs. In addition, reticular-cerebellar fibers conduct signals from the motor zone of the cerebral cortex, that is, they are the final segment of cortical-reticular-cerebellar connections.
The next “input” of afferent signals into the cerebellum is the olivocerebellar tract, which enters along the inferior cerebellar peduncles, crosses over, and ends on the Purkinje cells of the cerebellar cortex. The olivary nucleus is considered one of the most important “precerebellar” nuclei. It is believed that through the olivary nucleus and the olivo-cerebellar tract, the cerebellum receives signals from the cerebral cortex, the extrapyramidal system and proprioceptive information from the segmental apparatus of the spinal cord.
Along the pathways running from the vestibular nuclei (vestibular-cerebellar fibers) through the lower cerebellar peduncles to its most ancient part, the “flocculo-nodular system”, the cerebellum receives information from the receptors of the labyrinth, that is, information about acceleration forces, about the position of the head in space .
The main connections between the cerebral cortex and the cerebellum are through the pontine nuclei. Cortical-pontine pathways are directed to them from the cortex of all lobes of the hemispheres. In addition, collaterals branch off from the pyramidal tracts passing at the base of the bridge between its own nuclei to the cells of the nuclei. The axons of the cells of the own nuclei of the bridge undergo decussation and form the pontocerebellar tract, which is anatomically represented by the middle cerebellar peduncles; its fibers end on the cells of the cortex of the cerebellar hemispheres.
Having received information through its afferent pathways about the condition and tone of muscles, about the position of body parts, about the action of a number of environmental factors, as well as about the intended motor act, the cerebellum processes it and accordingly corrects the intended movement so that an optimal, maximally coordinated motor act is formed. Such coordination is achieved thanks to signals traveling from the cerebellum along its efferent connections to the motor centers of the brain and spinal cord.
The main integrative structures of the cerebellum are piriform neurons (Purkinje cells), which give rise to efferent fibers of the cerebellar cortex and are conventionally accepted as the third neuron of the cerebellar pathways. The axons of the piriform neurons go to the cerebellar nuclei, where the IV neuron of the cerebellar pathways is located. The projection efferent cerebellonuclear path begins from the tent nucleus; the fibers of this path pass in the inferior cerebellar peduncle and end in the motor nuclei of the cranial nerves and the nuclei of the reticular formation.
The most important efferent pathways of the cerebellum originate from the dentate nucleus. Serrated-red nuclear tract (tractus dentatorubralis)
passes in the superior cerebellar peduncle and, crossing, ends in the red nucleus, from where the red nucleus-spinal tract originates, which, along with the reticular-spinal cord, is the main descending pathway of the extrapyramidal system (the course of these pathways is described in the section on extrapyramidal tracts). These pathways end on the motor cells of the spinal cord. Thus, afferent and efferent cerebellar pathways provide regulation of movements at the level of the brainstem.
The transmission of nerve impulses from the cerebellum to the overlying floors of the brain occurs along the dentate-thalamic tract (tractus dentatothalamicus)
, which, like the dentate-red nuclear pathway, begins in the dentate nucleus, where the IV neuron is localized.
The cerebellar-thalamic tract passes in the superior cerebellar peduncles, crosses in the midbrain tegmentum (decussatio pedunculorum cerebellarium superiorum)
and approaches the inferolateral and central nuclei of the thalamus (V neuron). From the inferolateral nuclei, impulses are transmitted along the thalamo-cortical pathway to the cortex of the precentral gyrus, where the pyramidal tract originates.
The central nucleus of the thalamus transmits impulses coming from the cerebellum to the striatum, which belongs to the extrapyramidal system. Thus, through the dentate-thalamic, thalamo-cortical and thalamo-striatal pathways, the corrective influences of the cerebellum are transmitted to the motor pyramidal and extrapyramidal systems.
In addition, the cerebellum sends its effector impulses to the segmental apparatus of the spinal cord through the vestibular nuclei, as well as through the reticular formation, respectively, along the vestibular-spinal and reticular-spinal tracts. They, together with the red nuclear spinal tract, form the descending connections of the cerebellum.
4. The choroid of the eye, its parts. Accommodation mechanism.
The choroid of the eyeball, tunica vasculosa bulbi,
rich in blood vessels and pigment. It is directly adjacent on the inner side to the sclera, with which it is firmly fused at the point where the optic nerve exits the eyeball and at the border of the sclera with the cornea. The choroid is divided into three parts: the choroid itself, the ciliary body and the iris.
The choroid proper, choroidea,
It lines the large posterior part of the sclera, with which, in addition to the indicated places, it is loosely fused, limiting from the inside the so-called
perivascular space, spatium perichoroideale, existing between the membranes.
Ciliary body, corpus ciliare,
is a middle thickened section of the choroid, located in the form of a circular ridge in the area of the transition of the cornea to the sclera, behind the iris.
The ciliary body is fused with the outer ciliary edge of the iris. The posterior part of the ciliary body - the ciliary circle, orbiculus ciliaris,
has the appearance of a thickened circular stripe, passes into the choroid itself.
The anterior part of the ciliary body forms ciliary processes, processus ciliares.
These processes consist mainly of blood vessels and make up the ciliary
In the thickness of the ciliary body lies the ciliary muscle, m. ciliaris.
When the muscle contracts, accommodation of the eye occurs - adaptation to a clear vision of objects located at different distances.
In the ciliary muscle, meridional, circular and radial bundles of non-striated muscle cells are distinguished. The meridional (longitudinal) fibers
of this muscle originate from the edge of the cornea and from the sclera and are woven into the anterior part of the choroid itself.
When they contract, the shell moves anteriorly, as a result of which the tension of the ciliary girdle, zonula ciliaris,
on which the lens is attached, decreases.
At the same time, the lens capsule relaxes, the lens changes its curvature, becomes more convex, and its refractive power increases. Circular fibers, fibrae circulares,
narrow the ciliary body, bringing it closer to the lens, which also helps to relax the lens capsule.
Radial fibers, librae radiates,
start from the cornea and sclera in the region of the iridocorneal angle, are located between the meridional and circular bundles of the ciliary muscle, bringing these bundles closer together during their contraction. Elastic fibers present in the thickness of the ciliary body straighten the ciliary body when its muscle relaxes.
The iris, ins, is the most anterior part of the choroid, visible through the transparent cornea. It looks like a disk. In the center of the iris there is a round hole - the pupil, ririlla.
The diameter of the pupil is not constant: the pupil narrows in strong light and expands in the dark, acting as the diaphragm of the eyeball. The anterior surface of the iris faces the anterior chamber of the eyeball, and the posterior surface faces the posterior chamber and the lens.
Blood vessels are located in the connective tissue stroma of the iris. The cells of the posterior epithelium are rich in pigment, the amount of which determines the color of the iris (eye). In the thickness of the iris there are two muscles. Around the pupil there are circularly located bundles of smooth muscle cells - the pupillary sphincter, m. sphincter pupitlae,
and radially from the ciliary edge of the iris to its pupillary edge, thin bundles
of the muscle that dilate the pupil, i.e. dilatator puplllae
(pupil dilator), extend.
Ticket 7
Muscles and fascia of the neck...
Neck (cervix) -
part of the body connecting the head to the torso.
The lower border of the neck runs from the jugular notch of the sternum along the collarbone to the acromion of the scapula, then to the spinous process of the VII cervical vertebra. The upper border is a line from the mental protrusion along the base of the body of the lower jaw, along the posterior edge of the ramus to the temporomandibular joint, then to the base of the mastoid process of the temporal bone, along the superior nuchal line to the external occipital protrusion.
In the neck, two sections are conventionally distinguished: posterior
-
the neck (nucha),
the muscles of which are described in the section “Back”, and
the anterior one
-
the neck itself (collum).
In the neck itself there are 3 muscle groups:
superficial, middle
and
deep.
All neck muscles are paired.
Superficial neck muscles
1. Subcutaneous muscle of the neck (platysma) -
a wide thin plate lying on the anterolateral surface of the neck under the skin (Fig. 59). It starts from the pectoral fascia at the level of the second rib and from the deltoid fascia, attaches to the base of the body of the lower jaw, one part of the bundles is woven into the muscles in the area of the corner of the mouth, the other into the parotid-masticatory fascia.
Function: pulls back the skin of the neck and lowers the corner of the mouth. Innervation: facial nerve.
2. Sternocleidomastoid muscle (sternocleidomastoideus )
begins with two heads:
medial
- from the manubrium of the sternum and
lateral -
from the sternal end of the clavicle (Fig. 60, see Fig. 59). The heads, moving upward and backward, connect and attach to the mastoid process and the superior nuchal line of the occipital bone.
Function: with unilateral contraction, tilts the head in its direction, at the same time turning the head in the opposite direction. During bilateral contraction, he throws his head back.
Innervation: accessory nerve, Sp.
⇐ Previous2Next ⇒
Recommended pages:
Use the site search:
Flexig's posterior spinocerebellar tract
Named after the German neurologist and histologist PE Flechig (1847-1929), two-neuron. The 1st neuron is in the spinal ganglion. The dendrites end in the periosteum. The axons end on the neurons of the thoracic nucleus (neuron 2). Its axons form the posterior spinocerebellar tract. The path passes in the lateral funiculus and does not cross until the bark of the worm.
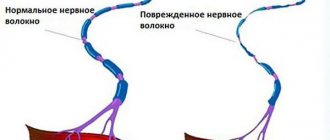
The proprioceptive pathways of the cerebellar direction and those associated with the cerebral cortex enable a person (especially athletes) to subtly feel their body and accurately perform highly differentiated movements. Processing various physical exercises is accompanied, first of all, by training the nervous system, and then the motor system.
TICKET No. 5
1. The significance of the works of V.P. Vorobyov, V.N. Tonkov, G.M. Iosifova, D.A. Zhdanov in anatomy.
2. Cecum and vermiform appendix. Projection onto the anterior abdominal wall, topography, peritoneal coverage.
3. Brain stem. Distribution of gray and white matter. Functions.
The significance of the works of V.P. Vorobyov, V.N. Tonkov, G.M. Iosifova, D.A. Zhdanov in anatomy.
An outstanding representative of the Kharkov school of anatomists was Vladimir Petrovich Vorobyov (1876-1937). Researcher of the autonomic nervous system, author of a method for studying nerves. Vorobyov described the nerve plexuses of the human heart and stomach, and was one of the first to study innervation using the method of electrical nerve stimulation in animals. He created the 5-volume Atlas of Human Anatomy. Developed and performed embalming by V.I. Lenin.
The founder of the Leningrad School of Anatomists, Vladimir Nikolaevich Tonkov (1872-1954), successfully worked in the field of experimental anatomy; for many years he headed the Department of Anatomy of the Military Medical Academy. He studied collateral circulation, plasticity of blood vessels, and blood supply to nerves. He was the first to use X-rays to study the skeleton in 1896.
The founder of the school of Russian lymphologists is Gordey Maksimovich Iosifov (1870-1933). His work “The Human Lymphatic System” is widely known. His students were the leading lymphologists D.A. Zhdanov, M.A. Kurdyumov, V.N. Nadezhdin.
Dmitry Arkadyevich Zhdanov (1908-1933) was awarded the State Prize for the monograph “Surgical anatomy of the thoracic duct and the main lymphatic collectors and nodes of the body” (1945). In 1952, his monograph “General Anatomy and Physiology of the Lymphatic System” was published.
Cecum and vermiform appendix. Projection onto the anterior abdominal wall, topography, peritoneal coverage.
Caecum, caecum,
located in the right iliac fossa and represents the initial expanded part of the colon below the point where the ileum enters the colon.
The posterior surface of the cecum lies on the iliacus and psoas major muscles, and its anterior surface is adjacent to the anterior abdominal wall. The cecum is covered with peritoneum on all sides (intraperitoneal position), but does not have a mesentery. On the posteromedial surface of the intestine, the ribbons of the colon converge at one point below. At this point, the vermiform appendix (appendix), appendix vertniformis, comes off.
The vermiform appendix is covered with peritoneum on all sides (located intraperitoneally) and has a mesentery. Less commonly, the base of the appendix is projected onto the anterior abdominal wall at the border between the outer and middle thirds of the line connecting the right superior anterior iliac spine and the umbilicus. More often, the base of the appendix is projected on the border between the outer and middle thirds of the line connecting the right and left superior anterior iliac spines. Basically, the appendix is located in the right iliac fossa, but can be located above or below. The direction of the appendix can be descending, lateral or ascending. In an ascending position, the appendix is often located behind the cecum.
The transition from the ileum to the cecum is the ileocecal opening, ostium ileocaecale,
bounded above and below by two folds forming
the ileocecal valve, vulva ileocaecalis
.
In front and behind, the folds of the valve converge and form the frenulum frenulum valvae ileocaecalisj.
Somewhat below the ileocecal valve, on the inner surface of the cecum there is
an opening of the appendix, ostium appendicis vermiformis. Vessels and nerves of the colon. The branches of the superior mesenteric artery, namely the ileocolic artery and its branches, approach the cecum and the appendix.
The stem part of the brain. Distribution of gray and white matter. Functions.
The brainstem includes the medulla oblongata, pons, midbrain and thalamus of the diencephalon. The brain stem contains vital centers for the regulation of respiration, blood circulation, cardiovascular activity, autonomic functions, the activity of the endocrine glands, etc. In its structure, the brain stem differs significantly from the spinal cord. First of all, it loses its characteristic metamerism in structure and cannot be divided into segments similar to the spinal cord. The gray matter of the trunk is represented by a cluster of nerve cell bodies or nuclei. It is separated by bundles of white matter.
Medulla
Medulla oblongata, myelencephalon, medulla oblongata
lies at the basis of GM, being a continuation of SM. In this regard, it combines the structural features of the SM and the initial section of the GM. The shape of the medulla oblongata resembles a truncated cone or onion, bulbus cerebri. Its length is approximately 25-30 mm, width at the base - 10 mm, at the top - 24 mm. Its lower border is the exit point of the first pair of spinal nerves. Above the medulla oblongata there is the pons, which from the ventral side looks like a constriction through the brain stem. The medulla oblongata, together with the pons and cerebellum, makes up the hindbrain, the cavity of which is the fourth cerebral ventricle.
Let's look at the ventral surface of the medulla oblongata. The anterior median fissure divides it into two symmetrical halves, and several grooves separate various structures from each other. The IX-XII pairs of cranial nerves depart from the medulla oblongata. VI - VIII pairs emerge from the groove separating the medulla oblongata from the pons. On the sides of the median fissure there are two longitudinal strands of white matter - pyramids.
These are fibers of the pyramidal tract coming from the cerebral cortex to the SC.
At the border with the SC, most of the fibers of this tract intersect, forming the decussatio pyramidum.
Lateral to the pyramids there are oval elevations - olives.
In their depths there is gray matter - the nuclei of olives. The olives have connections with the superior olivary nucleus of the pons, with the vestibular nuclei, and with the dentate nucleus of the cerebellum. The olives, together with the cerebellum, are involved in maintaining posture and motor learning.
Let us now consider the dorsal side of the medulla oblongata. Here it is divided into two symmetrical halves by the posterior median sulcus. On either side of it lie two bundles—tender (more medial) and wedge-shaped (more lateral). This is a continuation of the paths of the same name ascending from the SM. On the sides of the diamond-shaped fossa, thickenings are visible on the bundles - tubercles. Beneath them lie the tender and wedge-shaped nuclei. The cells of these nuclei are the second neurons of the pathways of the posterior cords of the spinal cord. These are the pathways of proprioceptive sensitivity of the cortical direction. The axons of these neurons travel along the posterior surface of the brain stem to the thalamus and are called the “medial lemniscus.” On the posterior surface of the medulla oblongata, the fibers of the medial lemniscus go to the opposite side, forming the decussation of the decussatio lemniscorum loop. Formation of the medial loop:
The posterior surface of the medulla oblongata forms the lower part of the rhomboid fossa.
The internal structure of the PM is represented by gray and white matter:
White matter occupies a large volume. It includes the so-called transit paths, i.e. ascending and descending tracts passing through the medulla oblongata without interruption (without forming synapses on its neurons).
Thus, the medulla oblongata performs reflex and conduction functions. The conductor function is that ascending and descending pathways pass through the brain stem (including the medulla oblongata), connecting the overlying parts of the brain, up to the cerebral cortex, with the spinal cord.
The reflex function is associated with the nuclei of the brain stem, through which reflex arcs are closed. It should be noted that in the medulla oblongata (mainly in the reticular nuclei) there are many vital centers - respiratory, vasomotor, centers of food reflexes (salivary, swallowing, chewing, sucking), centers of protective reflexes (sneezing, coughing, vomiting), etc. Therefore, damage to the medulla oblongata (trauma, swelling, hemorrhage, tumors) usually leads to very serious consequences.
Pons
Varoliev Bridge, pons Varolii
From the ventral side it looks like a thick ridge, bounded below by the medulla oblongata, and above by the cerebral peduncles (midbrain). The lateral border of the bridge is conventionally considered to be the exit of the trigeminal nerve. To the side of it are the middle cerebellar peduncles - fibers connecting the pons and the cerebellum. The ventral side is divided into two halves by the basilar groove, in which the basilar artery lies. Bridge length - 20 - 30 mm, width - 30 - 36 mm. The dorsal surface of the pons is the upper half of the rhomboid fossa - the bottom of the fourth ventricle.
Dorsal part of the bridge - tire
- consists mainly of gray matter, the lower -
the base
- of white. The tegmentum is separated from the base by fibers of the trapezoid body, coming from the ventral auditory nucleus and passing to the other side of the pons, i.e. related to the auditory pathway.
The gray matter of the pons, as well as in the medulla oblongata, is represented by nuclei:
trigeminal, abducens, facial, vestibulo-auditory nerves;
reticular formation;
3. nuclei of the superior olive and nuclei of the trapezoid body, the afferents of which are the ventral auditory nuclei;
4. locus coeruleus - a nucleus of noradrenergic (producing the neurotransmitter norepinephrine) neurons pigmented with melanin, which is why it got its name. This nucleus is involved in controlling the overall level of activation of the central nervous system, mainly in the direction of its increase, and is also associated with the regulation of emotions, memory, and autonomic functions
the bridge's own cores lie at its base. This is a group of nuclei that switch signals from the cerebral cortex to the cerebellum, i.e. fibers from the neurons of the cerebral cortex end on the neurons of the pontine nuclei, and axons from the pontine nuclei pass to the other side and enter the cerebellum, forming the middle cerebellar peduncles.
It should be noted that the bridge as a separate structure is present only in mammals. Its appearance is closely related to the development of the cerebral cortex, which is fully formed only in these animals.
White matter occupies the main volume of the bridge. Just as in the medulla oblongata, it includes transit pathways (pyramidal, rubrospinal, tectospinal, spinothalamic, etc.). Separately, it is necessary to note the cortico-pontine-cerebellar tract - fibers going from the cerebral cortex to the cerebellum with switching on the own nuclei of the bridge.
The main auditory pathway, the lateral lemniscus (lateral lemniscus), is also formed in the pons. It is a mass of crossed and uncrossed axons from various auditory nuclei, namely the superior olivary nucleus, the posterior cochlear nucleus and the trapezius nucleus. The lateral lemniscus ends in the subcortical hearing centers: in the inferior colliculi of the midbrain quadrigeminal and in the medial geniculate body.
Midbrain
Midbrain, mesencephalon,
the smallest section of the brain, its length is approximately 2 cm. The cavity of the midbrain - the cerebral (Sylvian) aqueduct has a diameter of about 1 mm. Two pairs of cranial nerves emerge from the midbrain - the oculomotor nerve (III pair) and the trochlear nerve (IV pair). Let us recall that the trochlear nerve leaves the brain on the dorsal side, then goes around the cerebral peduncles and passes to the ventral side.
On the dorsal side of the midbrain there is a roof, tectum,
consisting of two pairs of tubercles - the lower and upper colliculi of the quadrigeminal. They are separated by mutually perpendicular grooves. The handle of the colliculus extends from each tubercle - fibers connecting the superior colliculus to the lateral geniculate body of the metathalamus; the inferior colliculus connects to the medial geniculate body. Upper colliculi and lats. the geniculate bodies are the subcortical centers of vision. The motor pathways of the fibers of the tectospinal tract, the fibers going to the nuclei of the oculomotor nerves, also depart from them. These connections contribute to the main function of the superior colliculi - organizing movements in response to a new stimulus (turning the head, eyes, ears towards the stimulus). This innate reaction is called the orienting reflex.
In phylogeny, the inferior colliculi appear only in mammals and are auditory centers. The auditory fibers of the lateral lemniscus end here.
The cerebral peduncles are located on the ventral side. They emerge from the bridge, head forward and, gradually diverging to the sides, plunge into the thickness of the cerebral hemispheres. Between the legs lies the interpeduncular fossa, in the bottom of which there are many small holes through which blood vessels pass. This area is called the posterior perforated substance.
The cerebral peduncles are divided into the tegmentum,
and the underlying basis, basis. The border between them is the black substance substantia nigra.
The roof of the brain consists of gray matter, the base is made of white matter (descending tracts only), and in the tegmentum, among the fibers of the white matter, lie the nuclei of gray matter.
TICKET No. 6
1. Development of anatomy in the 19th century (P.A. Zagorsky, D.N. Zernov, N.I. Pirogov, P.F. Lesgaft).
2. Salivary glands: topography, structure, excretory ducts, blood supply and innervation.
3. Pyramid tracts. Topography of pyramidal tracts in various parts of the brain.