В продолговатом мозге, аксоны вторых нейронов проходят между нижней оливой и нижней мозжечковой ножкой. В покрышке моста они приближаются к его дорсальной поверхности. На границе со средним мозгом волокна переднего спинно-мозжечкового пути поворачивают дорсально в область верхнего мозгового паруса. Здесь часть волокон еще раз переходит на противоположную сторону, а затем через верхние ножки мозжечка (pedunculi cerebellares superiores) достигают передневерхних отделов мозжечка и входят в ядро шатра, nucleus fastigii, червя мозжечка, vermis cerebelli. ). Ядро шатра является коллектором афферентной информации. Отсюда она поступает к грушевидным нейронам коры мозжечка — клеткам Пуркинье. Ян Евангелист Пуркинье, Jan Evangelista Purkinje, 1787-1869, чешский физиолог, гистолог, эмбриолог и фармаколог. Грушевидные нейроны обрабатывают полученную информацию и передают ее к зубчатому ядру (nucleus dentatus) одноименного полушария мозжечка. В этой структуре заканчиваются афферентные пути. Зубчатое ядро — конечное звено регуляторов мозжечка. Его нейроны формируют и посылают эфферентные (управляющие) сигналы через верхние мозжечковые ножки к красному ядру среднего мозга противоположной стороны (перекрест Вернекинга). Можно проследить системы волокон, по которым информация из коры червя достигает красного ядра, полушария мозжечка, а также вышележащих отделов мозга — коры полушарий большого мозга. Из коры червя через пробковидное и шаровидное ядра информация через верхнюю мозжечковую ножку направляется к красному ядру противоположной стороны (мозжечково-покрышечный путь). Кора червя связана ассоциативными волокнами с корой полушария мозжечка, откуда информация поступает в зубчатое ядро мозжечка. Существуют также связи мозжечка с корой, осуществляющиеся через таламус. В частности, аксоны нейронов зубчатого ядра через верхнюю мозжечковую ножку выходят в покрышку моста, переходят на противоположную сторону и направляются к таламусу. Переключившись в таламусе на следующий нейрон, пути следуют в кору большого мозга, в постцентральную извилину. Описанные спинно-мозжечковые пути основные, но не единственные известные спинно-мозжечковые пути. Частью заднего спинно-мозжечкового тракта считают клиновидно-мозжечковый тракт (tractus cuneocerebellaris). Частью переднего спинно-мозжечкового тракта считают ростральный спинно-мозжечковый тракт (tractus spinocerebellaris rostralis). Существуют также непрямые спинно-мозжечковые тракты. Среди них наиболее изучены: спинно-оливарно-мозжечковые тракты, спинно-бульбо-мозжечковые тракты и спинно-ретикуло-мозжечковые тракты.
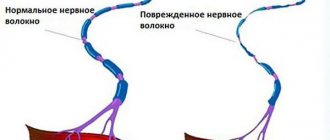
Благодаря информации, поступающей по передним спинно-мозжечковым путям и задним спинно-мозжечковым путям, мозжечок, как один из нервных центров в иерархии регуляторов опорно-двигательной системы, участвует в управлении движениями, обеспечивающими равновесие тела без участия высших отделов головного мозга (коры полушарий большого мозга), без участия сознания.
См.: Неврология: Словарь, Неврология: Ресурсы Интернет.
| БИБЛИОТЕКА = LIBRARY Физиология человека = Human Physiology, Анатомия человека = Human Anatomy, Биохимия человека = Human Biochemistry, Психология человека = Human Psychology, Медицина = Medicine, Математика = Mathematics, Химия = Chemistry, Физика = Physics, Общенаучная литература = General Science Lexis. Щелкни здесь и получи доступ к любому источнику библиотеки сайта! Click here and receive access to the any reference of the library! «Я У Ч Е Н Ы Й И Л И . . . Н Е Д О У Ч К А ?» Т Е С Т В А Ш Е Г О И Н Т Е Л Л Е К Т А Предпосылка : Эффективность развития любой отрасли знаний определяется степенью соответствия методологии познания — познаваемой сущности. |
Ошибка? Щелкни здесь и исправь ее! Поиск на сайте E-mail автора (author)
Пути проприоцептивной чув-ти мозжечкового направления (Флексига и Говерса)
⇐ ПредыдущаяСтр 2 из 36Следующая ⇒
Издавна считалось, что мозжечок является одним из центров координации и синергии движений, регуляции тонуса мускулатуры, поддержания равновесия. Академик Л.А.Орбели пришел к заключению, что «мозжечок является высшим адаптационно-трофическим центром», где происходит интеграция сомато-вегетативных функций.
В настоящее время мозжечок рассматривается как своеобразная система, координирующая двигательные акты. Благодаря своим обширным связям мозжечок получает информацию от рецепторов, находящихся в мышцах, сухожилиях, суставах, а также от вестибулярного, зрительного и слухового анализаторов, от ретикулярной формации. Этот огромный приток сенсорной информации служит в конечном итоге для регуляции двигательных функций. Интегрированная информация выдается в виде эфферентных сигналов к ведущим моторным центрам спинного и головного мозга.
По-видимому, мозжечок не играет ведущей роли в организации двигательных актов, не программирует движения, а оптимизирует программы движения по мере их осуществления. Мозжечок принимает участие в контроле движений, внося соответствующие коррективы в деятельность моторных центров головного и спинного мозга. Предполагается, что мозжечок не только регулирует активность отдельных двигательных центров, но и согласует их работу.
Следовательно, важнейшими афферентными связями мозжечка являются проприоцептивные пути от спинного мозга, вестибулярных ядер, экстрапирамидной системы, двигательных центров коры полушарий головного мозга; важнейшими эфферентными связями — пути к двигательным центрам головного и спинного мозга.
Мозжечок получает информацию о состоянии и деятельности мышечно-суставной системы по нескольким каналам. Это, во-первых, прямые спинно-мозжечковые тракты, непосредственно связывающие спинной мозг с мозжечком, и, во-вторых, проводники, переключающиеся на пути к мозжечку в ядрах ствола мозга. Эти ядра именуют «предмозжечковыми».
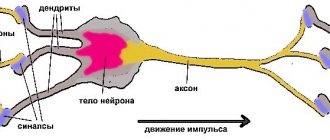
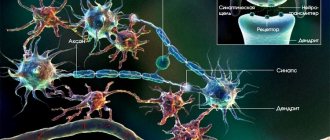
Проприоцепторы заложены в мышцах, сухожилиях, суставных поверхностях, суставных капсулах, связках, фасциях, надкостнице. Раздражения передаются по чувствительным волокнам спинномозговых нервов клеткам спинномозговых ганглиев (I нейрон) и от них по волокнам задних корешков в серое вещество спинного мозга. Отсюда берут начало два анатомически раздельных нервных пути:
А. Задний спинно-мозжечковый путь (tractus spinocerebellaris posterior)
(пучок Флексига) начинается от клеток грудного ядра
(nucleus thoracicus)
(II нейрон). Волокна этого пути проходят, не образуя перекреста, по периферии бокового канатика спинного мозга, продолжаются в продолговатый мозг и вступают в нижние мозжечковые ножки. Оканчивается путь в коре червя мозжечка своей стороны (paleocerebellum).
Б. Передний спинно-мозжечковый путь (tractus spinocerebellaris anterior)
(пучок Говерса) берет начало от клеток промежуточно-медиального ядра
(substantia intermedia medialis)
(II нейрон). Волокна пути преимущественно переходят на противоположную сторону в передней белой спайке спинного мозга (первый перекрест) и по периферии бокового канатика идут в восходящем направлении, проходят через продолговатый мозг и мост и достигают среднего мозга. Затем передний спинно-мозжечковый путь проходит по верхним мозжечковым ножкам, образуя второй (частичный) перекрест в переднем мозговом парусе, и аналогично заднему пути оканчивается в коре червя мозжечка.
Структурные различия двух спинно-мозжечковых путей предопределяют их функциональные различия. Считается, что по заднему пути мозжечок получает информацию от мышечных и сухожильных рецепторов каждой обособленной группы мышц. По переднему пути в мозжечок поступают сигналы, по-видимому, от больших мышечных групп.
Ко второму типу связей «опорно-двигательный аппарат — спинной мозг — мозжечок», переключающихся на ядрах ствола мозга, относятся задние наружные дугообразные волокна.
В составе нижних ножек к коре полушарий мозжечка проходят ретикулярно-мозжечковые волокна, которые замыкают спинно-ретикулярно-мозжечковые связи. Предполагается, что по ним мозжечок получает дополнительную проприоцептивную информацию о движениях, совершаемых скелетными мышцами конечностей. Кроме того, ретикулярно-мозжечковые волокна проводят сигналы от двигательной зоны коры полушарий головного мозга, то есть являются конечным отрезком корково-ретикулярно-мозжечковых связей.
Следующий «вход» афферентных сигналов в мозжечок — оливо-мозжечковый путь, вступающий по нижним мозжечковым ножкам, подвергающийся перекресту и заканчивающийся на клетках Пуркинье коры мозжечка. Оливное ядро рассматривают как одно из важнейших «предмозжечковых» ядер. Считается, что через оливное ядро и оливо-мозжечковый путь мозжечок получает сигналы от коры полушарий, экстрапирамидной системы и проприоцептивную информацию от сегментарного аппарата спинного мозга.
По проводящим путям, идущим от вестибулярных ядер (преддверно-мозжечковые волокна) через нижние ножки мозжечка к наиболее древней его части, «флокуло-нодулярной системе», мозжечок получает информацию от рецепторов лабиринта, то есть информацию о силах ускорения, о положении головы в пространстве.
Главные связи коры больших полушарий с мозжечком осуществляются через собственные ядра моста. К ним от коры всех долей полушарий направляются корково-мостовые пути. Кроме того, от пирамидных путей, проходящих в основании моста между его собственными ядрами, ответвляются коллатерали к клеткам ядер. Аксоны клеток собственных ядер моста подвергаются перекресту и образуют мосто-мозжечковый тракт, который анатомически представлен средними мозжечковыми ножками, его волокна заканчиваются на клетках коры полушарий мозжечка.
Получив по своим афферентным путям информацию о состоянии и тонусе мышц, о положении частей тела, о действии ряда факторов внешней среды, а также о предполагаемом двигательном акте, мозжечок перерабатывает ее и соответственно корригирует намечающееся движение так, чтобы сформировался оптимальный, максимально координированный двигательный акт. Подобная координация достигается благодаря сигналам, следующим от мозжечка по его эфферентным связям к двигательным центрам головного и спинного мозга.
Основным интегративными структурами мозжечка являются грушевидные нейроны (клетки Пуркинье), которые дают начало эфферентным волокнам коры мозжечка и условно принимаются за III нейрон мозжечковых проводящих путей. Аксоны грушевидных нейронов идут к ядрам мозжечка, где заложен IV нейрон мозжечковых путей. От ядра шатра начинается проекционный эфферентный мозжечково-ядерный путь, волокна этого пути проходят в нижней мозжечковой ножке и оканчиваются в двигательных ядрах черепных нервов и ядрах ретикулярной формации.
Наиболее важные эфферентные пути мозжечка берут начало от зубчатого ядра. Зубчато-красноядерный путь(tractus dentatorubralis)
проходит в верхней мозжечковой ножке и, перекрещиваясь, оканчивается в красном ядре, откуда берет начало красноядерно-спинномозговой путь, являющийся, наряду с ретикулярно-спинномозговым путем, главным нисходящим путем экстрапирамидной системы (ход этих путей описан в разделе, посвященном экстрапирамидным путям). Оканчиваются эти пути на двигательных клетках спинного мозга. Таким образом, афферентные и эфферентные мозжечковые пути обеспечивают регуляцию движений на уровне стволовой части головного мозга.
Передача нервных импульсов из мозжечка в вышележащие этажи мозга осуществляется по зубчато-таламическому пути (tractus dentatothalamicus)
, который, как и зубчато-красноядерный путь, начинается в зубчатом ядре, где локализуется IV нейрон. Мозжечково-таламический путь проходит в верхних мозжечковых ножках, перекрещивается в покрышке среднего мозга
(decussatio pedunculorum cerebellarium superiorum)
и подходит к нижнелатеральным и центральному ядрам таламуса (V нейрон). Из нижнелатеральных ядер импульсы передаются по таламо-корковому пути в кору предцентральной извилины, где берет начало пирамидный путь.
Центральное ядро таламуса передает импульсы, приходящие из мозжечка, полосатому телу, относящемуся к экстрапирамидной системе. Таким образом, за счет зубчато-таламического, таламо-коркового и таламо-стриального путей передаются корригирующие влияния мозжечка на двигательные пирамидную и экстрапирамидную системы.
Кроме того, мозжечок посылает свои эффекторные импульсы к сегментарному аппарату спинного мозга через вестибулярные ядра, а также через ретикулярную формацию, соответственно, по преддверно-спинномозговому и ретикулярно-спинномозговому путям. Они вместе с красноядерно-спинномозговым трактом образуют нисходящие связи мозжечка.
4. Сосудистая оболочка глаза, ее части. Механизм аккомодации.
Сосудистая оболочка глазного яблока, tunica vasculosa bulbi,
богата кровеносными сосудами и пигментом. Она непосредственно прилежит с внутренней стороны к склере, с которой прочно сращена у места выхода из глазного яблока зрительного нерва и у границы склеры с роговицей. В сосудистой оболочке выделяют три части: собственно сосудистую оболочку, ресничное тело и радужку.
Собственно сосудистая оболочка, choroidea,
выстилает большую заднюю часть склеры, с которой, кроме указанных мест, сращена рыхло, ограничивая изнутри имеющееся между оболочками так называемое
околососудистое пространство, spatium perichoroideale.
Ресничное тело, corpus ciliare,
представляет собой средний утолщенный отдел сосудистой оболочки, расположенный в виде кругового валика в области перехода роговицы в склеру, позади радужки. С наружным ресничным краем радужки ресничное тело сращено. Задняя часть ресничного тела —
ресничный кружок, orbiculus ciliaris,
имеет вид утолщенной циркулярной полоски, переходит в собственно сосудистую оболочку. Передняя часть ресничного тела образует
ресничные отростки, processus ciliares.
Эти отростки состоят в основном из кровеносных сосудов и составляют
ресничный венец, corona ciliaris.
В толще ресничного тела залегает ресничная мышца, m. ciliaris.
При сокращении мышцы происходит аккомодация глаза — приспособление к четкому видению предметов, находящихся на различном расстоянии. В ресничной мышце выделяют меридиональные, циркулярные и радиарные пучки не-исчерченных мышечных клеток.
Меридиональные (продольные) волокна,
этой мышцы берут начало от края роговицы и от склеры и вплетаются в переднюю часть собственно сосудистой оболочки. При их сокращении оболочка смещается кпереди, в результате чего уменьшается натяжение
ресничного пояска, zonula ciliaris,
на котором укреплен хрусталик. Капсула хрусталика при этом расслабляется, хрусталик изменяет свою кривизну, становится более выпуклым, а его преломляющая способность увеличивается.
Циркулярные волокна, fibrae circulares,
суживают цилиарное тело, приближая его к хрусталику, что также способствует расслаблению капсулы хрусталика.
Радиальные волокна, librae radiates,
начинаются от роговицы и склеры в области радужно-роговичного угла, располагаются между меридиональными и циркулярными пучками ресничной мышцы, сближая эти пучки при своем сокращении. Присутствующие в толще цилиарного тела эластические волокна расправляют цилиарное тело при расслаблении его мышцы.
Радужка, ins,— самая передняя часть сосудистой оболочки, видимая через прозрачную роговицу. Она имеет вид диска. В центре радужки имеется круглое отверстие — зрачок, рирillа.
Диаметр зрачка непостоянный: зрачок суживается при сильном освещении и расширяется в темноте, выполняя роль диафрагмы глазного яблока.Передняя поверхность радужки обращена в сторону передней камеры глазного яблока, а задняя — к задней камере и хрусталику.
В соединительнотканной строме радужки располагаются кровеносные сосуды. Клетки заднего эпителия богаты пигментом, от количества которого зависит цвет радужки (глаза). В толще радужки лежат две мышцы. Вокруг зрачка циркулярно расположены пучки гладких мышечных клеток — сфинктер зрачка, m. sphincter pupitlae,
а радиально от ресничного края радужки до ее зрачкового края простираются тонкие пучки
мышцы, расширяющей зрачок, т. dilatator puplllae
(расширитель зрачка).
Билет 7
Мышцы и фасции шеи…
Шея(cervix) —
часть тела, соединяющая голову с туловищем. Нижняя граница шеи проходит от яремной вырезки грудины по ключице до акромиона лопатки, далее — к остистому отростку VII шейного позвонка.
Верхняя граница — это линия от подбородочного выступа по основанию тела нижней челюсти, по заднему краю ветви к височно-нижнечелюстному суставу, далее к основанию сосцевидного отростка височной кости, по верхней выйной линии к наружному затылочному выступу.
В шее условно различают 2 отдела: задний
—
выю (nucha),
мышцы которого описаны в разделе «Спина», и
передний
—
собственно шею (collum).
В собственно шее выделяют 3 группы мышц:
поверхностные, средние
и
глубокие.
Все мышцы шеи парные.
Поверхностные мышцы шеи
1. Подкожная мышца шеи(platysma) —
широкая тонкая пластинка, залегающая на переднебоковой поверхности шеи под кожей (рис. 59). Начинается от грудной фасции на уровне второго ребра и от дельтовидной фасции, прикрепляется к основанию тела нижней челюсти, одной частью пучков вплетается в мышцы в области угла рта, другой — в околоушно-жевательную фасцию.
Функция: оттягивает кожу шеи и опускает угол рта. Иннервация: лицевой нерв.
2. Грудино-ключично-сосцевидная мышца(т. sternocleidomastoideus)
начинается двумя головками:
медиальной
— от рукоятки грудины и
латеральной —
от грудинного конца ключицы (рис. 60, см. рис. 59). Головки, направляясь вверх и назад, соединяются и прикрепляются к сосцевидному отростку и верхней выйной линии затылочной кости.
Функция: при одностороннем сокращении наклоняет голову в свою сторону, одновременно поворачивает голову в противоположную сторону. При двустороннем сокращении запрокидывает голову.
Иннервация: добавочный нерв, Сп.
⇐ Предыдущая2Следующая ⇒
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
Задний спинно-мозжечковый путь Флексига
Назван в честь немецкого невропатолога и гистолога P.E. Flechig (1847-1929), двухнейронный. 1-ый нейрон – в спинальном ганглии. Дендриты оканчиваются в надкостнице. Аксоны оканчиваются на нейронах грудного ядра (2 нейрон). Его аксоны образуют задний спинно-мозжечковый путь. Путь проходит в боковом канатике, не перекрещивается до коры червя.
Проприоцептивные пути мозжечкового направления и связанные с корой больших полушарий дают возможность человеку (особенно спортсменам) тонко чувствовать свое тело и точно выполнять высокодифференцированные движения. Обработка различных физических упражнений сопровождается в первую очередь, тренировкой нервной системы, а затем уже двигательного аппарата.
БИЛЕТ № 5
1. Значение работ В.П.Воробьева, В.Н.Тонкова, Г.М. Иосифова, Д.А. Жданова в анатомии.
2. Слепая кишка и червеобразный отросток. Проекция на переднюю брюшную стенку топография, покрытие брюшиной.
3. Стволовая часть головного мозга. Распределение серого и белого веществ. Функции.
Значение работ В.П.Воробьева, В.Н.Тонкова, Г.М. Иосифова, Д.А. Жданова в анатомии.
Выдающимся представителем Харьковской школы анатомов был Владимир Петрович Воробьев (1876-1937). Исследователь вегетативной нервной системы, автор метода изучения нервов. Воробьев описал нервные сплетения сердца и желудка человека, одним из первых начал изучение иннервации методом электростимуляции нервов у животных. Он создал 5 томный «Атлас анатомии человека». Разработал и выполнил бальзамирование В.И. Ленина.
В области экспериментальной анатомии успешно работал основатель Ленинградской школы анатомов Владимир Николаевич Тонков (1872-1954) в течение многих лет возглавлял кафедру анатомии Военно-медицинской академии. Он исследовал коллатеральное кровообращение, пластичность кровеносных сосудов, кровоснабжение нервов. Первым в 1896 г. применил рентгеновские лучи для изучения скелета.
Основоположник школы русских лимфологов является Гордей Максимович Иосифов (1870-1933). Широко известен его труд «Лимфатическая система человека». Его учениками явились крупнейшие лимфологи Д.А.Жданов, М.А.Курдюмов, В.Н.Надеждин.
Дмитрий Аркадьевич Жданов (1908-1933) за монографию «Хирургическая анатомия грудного протока и главных лимфатических коллекторов и узлов туловища» (1945) удостоен Государственной премии. В 1952 г. вышла его монография «Общая анатомия и физиология лимфатической системы».
Слепая кишка и червеобразный отросток. Проекция на переднюю брюшную стенку топография, покрытие брюшиной.
Слепая кишка,caecum,
расположена в правой подвздошной ямке и представляет собой начальную расширенную часть толстой кишки ниже места впадения подвздошной кишки в толстую. Задней поверхностью слепая кишка лежит на подвздошной и большой поясничной мышцах, а передняя ее поверхность прилежит к передней брюшной стенке. Брюшиной слепая кишка покрыта со всех сторон (интраперитонеальное положение), однако брыжейки не имеет. На заднемедиальной поверхности кишки внизу сходятся в одной точке ленты ободочной кишки. В этом месте отходит
червеобразный отросток (аппендикс), appendix vertniformis.
Червеобразный отросток покрыт брюшиной со всех сторон (расположен интраперитонеально) и имеет брыжейку. Реже основание червеобразного отростка проецируется на переднюю брюшную стенку на границе между наружной и средней третями линии, соединяющей правую верхнюю переднюю подвздошную ость и пупок. Чаще основание червеобразного отростка проецируется на границе между наружной и средней третями линии, соединяющей правую и левую верхние передние подвздошные ости В основном червеобразный отросток расположен в правой подвздошной ямке, но может находиться выше или ниже. Направление червеобразного отростка может быть нисходящим, латеральным или восходящим. При восходящем положении червеобразный отросток нередко располагается позади слепой кишки.
Переходом подвздошной кишки в слепую является илеоцекальное отверстие,ostium ileocaecale,
ограниченное сверху и снизу двумя складками, образующими
илеоцекальный клапан,vulva ileocaecalis
. Спереди и сзади складки клапана сходятся и образуют
уздечку илеоцекального клапана,frenulum valvae ileocaecalisj.
Несколько ниже илеоцекального клапана на внутренней поверхности слепой кишки имеется
отверстие червеобразного отростка (аппендикса),ostium appendicis vermiformis.Сосуды и нервы ободочной кишки.К слепой кишке и червеобразному отростку подходят ветви верхней брыжеечной артерии, а именно подвздошно-ободочная артерия с ее ветвями.
Стволовая часть головного мозга. Распределение серого и белого веществ. Функции.
К стволу мозга относятся: продолговатый мозг, мост, средний мозг и таламусы промежуточного мозга. В мозговом стволе находятся жизненно важные центры регуляции дыхания, кровообращения, сердечно-сосудистой деятельности, вегетативных функций, деятельности эндокринных желез и др. По своему строению ствол мозга значительно отличается от спинного мозга. Прежде всего, он утрачивает характерную метамерность в строении и его нельзя разделить на сегменты, подобные спинно-мозговым. Серое вещество ствола представлено скоплением тел нервных клеток или ядер. Оно разделяется пучками белого вещества.
Продолговатый мозг
Продолговатый мозг, myelencephalon, medulla oblongata
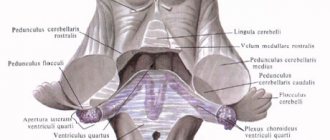
лежит в основании ГМ, являясь продолжением СМ. В связи с этим он сочетает черты строения СМ и начального отдела ГМ. По форме продолговатый мозг напоминает усеченный конус или луковицу, bulbus cerebri. Длина его примерно 25-30 мм, ширина в основании — 10 мм, у вершины — 24 мм. Его нижняя граница — место выхода I пары спинномозговых нервов. Выше продолговатого мозга расположен варолиев мост, внешне представляющий собой с брюшной стороны как будто перетяжку через ствол мозга. Продолговатый мозг вместе с варолиевым мостом и мозжечком составляет задний мозг, полостью которого является IV мозговой желудочек.
Рассмотрим вентральную поверхность продолговатого мозга. Передняя срединная щель делит его на две симметричные половины, а несколько борозд отделяют друг от друга различные структуры. От продолговатого мозга отходят IX — XII пары черепных нервов. VI — VIII пары, выходят из борозды, отделяющей продолговатый мозг от моста. По бокам от срединной щели находятся два продольных тяжа белого вещества — пирамиды.
Это волокна пирамидного тракта, идущего из коры больших полушарий в СМ.
На границе со СМ большинство волокон этого тракта перекрещиваются, образуя перекрест пирамид decussatio pyramidum.
Латерально от пирамид лежат овальные возвышения — оливы.
В их глубине находится серое вещество — ядра олив. Оливы имеют связь с верхним оливным ядром моста, с вестибулярными ядрами, с зубчатым ядром мозжечка. Оливы вместе с мозжечком принимают участие в поддержании позы и двигательном обучении.
Рассмотрим теперь дорсальную сторону продолговатого мозга. Здесь он разделен на две симметричных половины задней срединной бороздой. По бокам от нее лежат два пучка— нежный (более медиальный) и клиновидный (более латеральный). Это продолжение одноименных путей, восходящих из СМ. По бокам от ромбовидной ямки на пучках видны утолщения — бугорки. Под ними лежат нежное и клиновидное ядра. Клетки этих ядер являются вторыми нейронами проводящих путей задних канатиков спинного мозга. Это пути проприоцептивной чувствительности коркового направления. Аксоны этих нейронов идут по задней поверхности ствола мозга до таламуса и называются «медиальная петля». На задней поверхности продолговатого мозга волокна медиальной петли идут на противоположную сторону, образуя перекрест петли decussatio lemniscorum. Формирование медиальной петли:
Задняя поверхность продолговатого мозга образует нижний отдел ромбовидной ямки.
Внутреннее строение ПМ представлено серым веществом и белым:
Белое веществозанимает большой объем. Оно включает так называемые транзитные тракты, т.е. восходящие и нисходящие тракты, проходящие через продолговатый мозг не прерываясь (не образуя синапсов на его нейронах).
Таким образом, продолговатый мозг выполняет рефлекторную и проводниковую функции. Проводниковая функция заключается в том, что через ствол мозга (в том числе и через продолговатый мозг) проходят восходящие и нисходящие пути, связывающие вышележащие отделы мозга, вплоть до коры больших полушарий, со спинным мозгом.
Рефлекторная функция связана с ядрами мозгового ствола, через которые замыкаются рефлекторные дуги. Надо отметить, что в продолговатом мозгу (в основном, в ретикулярных ядрах) находятся многие жизненно важные центры— дыхательный, сосудодвигательный, центры пищевых рефлексов (слюнных, глотательных, жевательных, сосательных), центры защитных рефлексов (чихания, кашля, рвоты) и др. Поэтому повреждения продолговатого мозга (травма, отек, кровоизлияние, опухоли) обычно приводят к очень тяжелым последствиям.
Варолиев мост
Варолиев мост, pons Varolii
выглядит с вентральной стороны как толстый валик, ограниченный снизу продолговатым мозгом, а сверху ножками мозга (средний мозг). Латеральной границей моста условно считается выход тройничного нерва. Сбоку от него находятся средние ножки мозжечка — волокна, связывающие мост и мозжечок. Вентральную сторону делит на две половины базилярная борозда, в которой лежит базилярная артерия. Длина моста — 20 — 30 мм, ширина — 30 — 36 мм. Дорсальная поверхность моста представляет собой верхнюю половину ромбовидной ямки — дна IV желудочка.
Дорсальная часть моста — покрышка
— состоит в основном из серого вещества, нижняя —
основание
— из белого. Покрышка отделена от основания волокнами трапециевидного тела, идущими от вентрального слухового ядра и переходящими на другую сторону моста, т.е. имеющими отношение к слуховому пути.
Серое веществомоста, так же как и в продолговатом мозгу, представлено ядрами:
тройничного, отводящего, лицевого, вестибуло-слухового нервов;
ретикулярной формации;
3. ядра верхней оливы и ядра трапециевидного тела, афферентами которых являются вентральные слуховые ядра;
4. голубое пятно — ядро из норадренергических (вырабытывающих медиатор норадреналин) нейронов, пигментированных меланином, за что оно и получило свое название. Это ядро участвует в управлении общим уровнем активации ЦНС, в основном, в сторону ее повышения, а также связано с регуляцией эмоций, памяти, вегетативных функций
собственные ядра моста — лежат в его основании. Это группа ядер, переключающих сигналы от коры больших полушарий к мозжечку, т.е. волокна от нейронов коры больших полушарий заканчиваются на нейронах ядер моста, а аксоны от ядер моста переходят на другую сторону и входят в мозжечок, образуя средние ножки мозжечка.
Надо отметить, что мост как отдельная структура присутствует только у млекопитающих. Его появление тесно связано с развитием коры больших полушарий, которая в полной мере формируется только у этих животных.
Белое веществозанимает основной объем моста. Так же, как и в продолговатом мозгу, оно включает транзитные пути (пирамидный, рубро-спинальный, текто-спинальный, спинно-таламический и т.д.). Отдельно надо отметить кортико-мосто-мозжечковой тракт — волокна, идущие от коры больших полушарий к мозжечку с переключением на собственных ядрах моста.
В мосту формируется также основной слуховой путь — латеральная петля (латеральный лемниск). Это масса перекрещенных и не перекрещенных аксонов от различных слуховых ядер, а именно верхнего оливного ядра, заднего улиткового и ядер трапециевидного тела. Заканчивается латеральный лемниск в подкорковых центрах слуха: в нижних холмиках четверохолмия среднего мозга и в медиальном коленчатом теле.
Средний мозг
Средний мозг,mesencephalon,
самый маленький отдел головного мозга, его длина примерно 2 см. Полость среднего мозга — мозговой (сильвиев) водопровод имеет диаметр около 1 мм. Из среднего мозга выходят две пары черепных нервов — глазодвигательный (III пара) и блоковый (IV пара). Напомним, что блоковой нерв выходит из мозга на дорсальной стороне, затем огибает ножки мозга и переходит на брюшную сторону.
На дорсальной стороне среднего мозга находится крыша, tectum,
состоящая из двух пар бугорков — нижних и верхних холмиков четверохолмия. Они разделены взаимно перпендикулярными бороздами. От каждого бугорка отходит ручка холмика — волокна, соединяющие верхний бугорок с латеральным коленчатым телом метаталамуса, нижний холмик соединяется с медиальным коленчатым телом. Верхние холмики и лат. коленчатые тела являются подкорковыми центрами зрения. От них также отходят двигательные пути волокна текто-спинального тракта, волокна, идущие к ядрам глазодвигательных нервов. Эти связи способствуют выполнению основной функции верхних холмиков — организации движений в ответ на новый раздражитель (поворот головы, глаз, ушных раковин в сторону стимула). Такую врожденную реакцию называют ориентировочным рефлексом.
Нижние холмики в филогенезе появляются только у млекопитающих и являются слуховыми центрами. Здесь заканчиваются — слуховые волокна латеральной петли.
На вентральной стороне расположены ножки мозга. Они выходят из моста, направляются вперед и, постепенно расходясь в стороны, погружаются в толщу больших полушарий. Между ножками лежит межножковая ямка, в дне которой есть множество мелких отверстий, через которые проходят кровеносные сосуды. Этот участок называется задним продырявленным веществом.
Ножки мозга разделены на покрышку, tegmentum,
и лежащее ниже основание, basis. Границей между ними служит черная субстанция substantia nigra.
Крыша мозга состоит из серого вещества, основание — из белого (только нисходящие тракты), в покрышке среди волокон белого вещества лежат ядра серого вещества.
БИЛЕТ № 6
1. Развитие анатомии в ХIХ веке (П.А. Загорский, Д.Н, Зернов, Н. И. Пирогов, П.Ф. Лесгафт).
2. Слюнные железы: топография, строение, выводные протоки, кровоснабжение и иннервация.
3. Пирамидные тракты. Топография пирамидных трактов в различных отделах мозга.