Structure of the hindbrain
The hindbrain consists of two main parts: the pons and the cerebellum located behind it. The first looks like a thick white cushion and is located above the medulla oblongata. The posterior dorsal surface of the pons is covered by the cerebellum, and the anterior ventral surface is represented by numerous transverse fibers that pass into the cerebellar middle peduncle. Nerve roots emerge into the bulbar-pontine groove. The main cerebral artery runs along the central sulcus of the pons. In general, the hindbrain has a rather complex structure.
Looking at the frontal section of the bridge, one can see both the large anterior and small posterior parts, delimited by a trapezoidal fibrous body. All interacting parts of the hindbrain provide conductive function. The cerebellum is otherwise called the small brain; it fills almost the entire space of the posterior cranial fossa. The usual weight of this organ is about 150 grams. The cerebral hemispheres are located above the cerebellum and are separated from it by a transverse fissure.
6.2.2. hindbrain
Our partners
rating of stucco molding sellers, orio
Youline.net - If you want to buy a sconce lamp and similar products, we offer you many items to choose from.
cheating of VK, VK surveys in Moscow
6. CENTRAL NERVOUS SYSTEM
6.2. Brain
The hindbrain develops from the postbrain vesicle ( tencephalon
), which is a derivative of the
rhombencephalon
. The ventral part of the hindbrain is a continuation of the brainstem structures and is called the pons. The pons contains the cavity of the hindbrain - part of the rhomboid fossa. The roof of the rhomboid fossa undergoes significant changes and develops into the cerebellum, which is the dorsal outgrowth of the hindbrain.
6.2.2.1. Pons
Varoliev Bridge ( pons Varolii
) is the ventral part of the hindbrain. The bridge itself forms the structures of the rostral part of the floor of the fourth ventricle. The dorsal surface of the pons is the superior triangle of the rhomboid fossa. The cavity of the rhomboid fossa narrows rostrally and passes into the midbrain aqueduct. From above, the cavity of the rhomboid fossa is covered by the superior medullary velum, which, together with the inferior medullary velum and the choroid plexus, forms the roof of the IV ventricle, which has the appearance of a tent. The lateral walls of the fourth ventricle in the area of the bridge are formed by the middle and superior cerebellar peduncles (see Fig. 25, 33).
The ventral surface of the pons is a powerful transverse fibrous convexity of white matter. A deep groove runs through the center of the ventral surface of the pons - the groove of the main artery of the brain ( suclus basil laris
). Laterally, the ventral convexity passes into the powerful middle cerebellar peduncles (see Fig. 25,
5
; 27,
4
).
Four pairs of cranial nerves depart from the pons (see Fig. 26; 27).
V - trigeminal nerve ( p. trigeminus
)
;
VI - abducens nerve ( p. abducens
)
;
VII - facial nerve ( p. facialis
)
;
VIII - prevestibulocochlearis, or auditory nerve ( p. vestibulo cochlearis
).
In cross sections, as in the medulla oblongata, white matter and gray matter nuclei are visible. The transverse fibers that make up the trapezoid body divide the thickness of the pons into larger ventral (base of the pons) and dorsal (tegmentum of the pons) parts. The ventral part is dominated by the white matter of the pathways, which are a continuation of the pathways of the midbrain peduncles. The gray matter of the ventral part of the pons forms its own pontine nuclei (nuclei of the base of the pons). In these nuclei, the descending corticomontine tracts and collaterals from the corticospinal tracts coming from the cerebral cortex end. Fibers extend from the pons' own nuclei, which pass to the opposite side and form trapezoidal bodies, which pass into the middle cerebellar peduncles.
The dorsal part of the pons is a direct continuation of the medulla oblongata. It contains the switching nuclei of the sensory systems, the nuclei of the cranial nerves and the reticular formation.
In phylogeny
In phylogeny
in lower vertebrates, the pons is not clearly separated from the medulla oblongata. It is isolated only in mammals. This occurs with the development of the cortex and the projection pathways descending from it. At the same time, the number of intrinsic nuclei of the ventral part of the pons increases. This determines the appearance and development of the middle cerebellar peduncles and its hemispheres. The ventral parts of the pons and the middle cerebellar peduncles are especially pronounced in humans.
In ontogeny
In ontogeny
The pons, as a part of the hindbrain, originates from the rhomboid medullary vesicle.
At the five-vesicle stage, the rhombencephalon divides into the accessory ( myelencephalon
), from which the medulla oblongata develops, and the hindbrain (
tencéphalon
). The roof of the hindbrain is transformed into the cerebellum, and its bottom and walls become pons structures. The cavity of the rhombencephalon remains common to the medulla oblongata and the pons and is the cavity of the fourth ventricle.
Almost all the nuclei of the cranial nerves of the pons originate in the rostral parts of the medulla oblongata. Their movement into the pons occurs after the formation of the medullary flexure. At the 7th week of embryonic development, cells of the pterygoid plate of the medulla oblongata migrate in the rostroventral direction and form a pontobulbar body on the ventral surface of the pons, which later turns into the pons' own nuclei.
6.2.2.2. Cerebellum
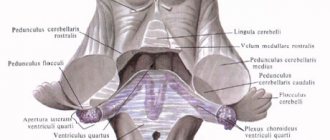
Rice. 29. Location of the cerebellum on the brain stem (part of the cerebellar tissue has been removed):
1 - superior cerebellar peduncle; 2 - middle cerebellar peduncle; 3 - leaf (cerebellar lobule); 4 - left hemisphere of the cerebellum; 5 - spinal cord; 6 - medulla oblongata; 7 - inferior cerebellar peduncle; 8 - bridge; 9 - midbrain
Cerebellum _
) is located on the dorsal surface of the brain stem.
The ventral surface of the cerebellum is adjacent to the sails of the IV ventricle and is closely connected with the stem structures by three pairs of cerebellar peduncles: with the medulla oblongata by the lower legs ( pedunculus cerebellaris inferior
) (Fig. 29,
7
), with the bridge by the middle legs (
pedunculus cerebellaris médius
) (Fig.
29
,
2
) and with the midbrain - the upper legs (
pedunculus cerebellaris superior
) (Fig. 29,
1
). The most powerful are the middle legs. All the legs emerge from the cerebellum side by side, and then the upper legs go to the midbrain along with the superior medullary velum, and the lower ones go to the medulla oblongata along with the inferior medullary velum. Rostral above the cerebellum lie the occipital lobes of the cerebrum, which extend beyond its dorsal boundaries of the cerebellum. The cerebellum is separated from the cerebrum by the deep transverse fissure of the brain. It, like the large brain, is covered with three membranes.
Anatomically, the human cerebellum consists of three main parts: two hemispheres ( hemi - spheria cerebelli
) and the middle part connecting them - the worm (
vermis cerebelli
). The surface of the cerebellum is indented with deep branching grooves. Deep grooves of the cerebellum divide the hemispheres and the vermis into lobules, which are combined into lobes: superior, posterior and inferior. The lobes are separated from each other by slits.
Between both hemispheres of the cerebellum there is a middle part cut by narrow, parallel grooves - the vermis.
(see Fig. 30a
,
13
)
. It has an upper surface, the upper worm, and a lower surface, the lower worm. Two longitudinal grooves on each surface of the cerebellum separate the superior and inferior vermis from the hemispheres
Rice. 30. Cerebellum:
bottom view (o): 1 - groove of the basilar artery; 2 - Varoliev Bridge; 3 - pyramid of the medulla oblongata; 4 - olive; 5 - worm nodule; 6 - choroid plexus of the IV ventricle; 7 - cerebellar amygdala; 8 - digastric lobe of the cerebellum; 9 - superior semilunar lobe of the cerebellum; 10 - horizontal sulcus of the cerebellum; 11 - lower semilunar lobule; 12 - gyri of the cerebellum; 13 - tubercle of the worm; 14 - worm pyramid; 15 - cerebellar valley; 16 - tongue of the worm; 17 — shred leg; 18 - scrap; 19 - trigeminal nerve; 20 - roots of the glossopharyngeal and vagus nerves; 21 - abducens nerve; right view (b): 1 - lateral geniculate body; 2 - medial geniculate body; 3 - optic tract; 4 - mamillary bodies; 5 - optic nerve; 6 - chiasma; 7 - funnel; 8 - pituitary gland; 9 - cerebral peduncle; 10 - lateral groove of the midbrain; 11 - trigeminal nerve; 12 — oblique beam of the bridge; 13 - abducens nerve; 14 - auditory and facial nerves; 15 - scrap; 16 - olive; 17 - hypoglossal nerve; 18 - external arcuate fibers; 19 - cerebellar amygdala; 20 - digastric lobule of the cerebellum; 21 - inferior semilunar lobe of the cerebellum; 22 - horizontal sulcus of the cerebellum; 23 - vagus nerve; 24 - glossopharyngeal nerve; 25 - superior semilunar lobule; 26 - quadrangular lobule; 27 - slope; 28 - top; 29 - groove of the trochlear nerve; 30 - lower tubercles of the quadrigeminal; 31 - lower arms of the quadrigeminal; 32 - upper tubercles of the quadrigeminal; 33.—upper handles of the quadrigeminal; 34 - cerebellar cushion. On the upper and lower worms, lobes are distinguished, which consist of several convolutions (Fig. 29, 3
).
Upper worm
from front to back consists of the following parts:
1) uvula of the cerebellum (Fig. 30 a
,
16
)
;
2
) central lobule (Fig. 33,
23
)
;
3) tubercle (Fig. 30a, 13
)
;
4) a leaf of the worm, in the form of a very narrow lobe located at the back, at the border of the transition of the upper worm to the lower one (Fig. 29, 3
).
On the bottom worm
, in the direction from front to back, the following lobes are distinguished:
1) nodule (Fig. 33, 20, to the anterior sections of which the posterior medullary velum is attached;
2) worm hub;
3) worm pyramid (Fig. 33, 16
).
On the superior surface of the cerebellar hemispheres
The following grooves and lobules are noted.
Quadrangular lobule ( lobulus quadrangularis
) (Fig. 30
b
,
26
) is divided by the anterior superior groove (
sulcus superior anterior
) into anterior and posterior parts.
The quadrangular lobule is limited by the posterior superior groove from the superior semilunar lobule ( lobulus semilunaris superior
) (Fig. 306,
25
).
Anterior to lobulus quadrangularis
small convolutions lie, the so-called wings of the central lobule.
Below and in front of the latter lie small areas of the cerebellum - the connections of the uvula ( vinculo lingulae
).
On the inferior surface of the cerebellar hemispheres
the following grooves and lobules are located.
A group of concentrically arranged convolutions forms the amygdala ( tonsilla
) (Fig. 30a,
7
; 30b
,
19
)
.
Outside and behind the tonsil lies the digastric lobule ( lo bulus bivénter
) (Fig. 30a,
8
, 30b
,
20
)
. The digastric lobule corresponds to the pyramid of the worm.
Outside the tonsil and in front of the digastric lobule is located the most anteriorly protruding part of the lower surface of the cerebellum - the flocculus
) (Fig. 30a,
18;
306,
15
).
Behind the digastric lobule lies the inferior semilunar lobule ( lobulus semilunaris inferior
)
(Fig. 30a,
11;
30b ,
21
).
The neural organization of the cerebellum differs significantly from that of brainstem structures. The bulk of neurons are concentrated on the surface and create the cerebellar cortex
(
cortex cerebelli
). Its area is large, since the bark is also present on the lateral surfaces of the furrows (about 80%).
Despite the fact that the mass of the cerebellum is only 1/9 of the mass of both cerebral hemispheres, the surface area of its cortex is equal to the surface area of one of them. The gray matter of the bark, located on the surface of the branching grooves, penetrates the white matter, like a tree. Therefore, the pattern formed by gray and white matter on sections of the cerebellum is called the cerebellar tree of life. Deep in the white matter there are accumulations of gray matter -
Rice. 31. Cerebellar nuclei:
1 - tent core; 2 - spherical nucleus; 3 - corky core; 4 - dentate core; 5 - cerebellar hemispheres; 6 - cerebellar vermis
Rice. 32. Scheme of the structure of the cerebellar cortex:
1 - stellate cell; 2 - Golgi cell; 3 - mossy fibers; 4 - granule cells; 5 - Purkinje cell
paired cerebellar nuclei
).
In the worm, on both sides of the midline, there are two tent nuclei ( nucleus fastigii cerebelli
) (Fig. 31,
1
), lateral to the tent nuclei in the cerebellar hemispheres we observe spherical nuclei (
nucleus globusus cerebelli
) (Fig. 31,
2
).
Lateral to the latter, in the hemispheres there are cork-shaped nuclei ( nucleus emboliformis
) (Fig. 31,
3
), and even further - the largest nuclei of the hemispheres, jagged (
nucleus dentatus
) (Fig. 31,
4
), which are a wave-like bending plate of gray matter.
The cerebellar cortex is clearly divided into three layers (Fig. 32):
1) outer - molecular layer
(
stratum moleculare
)
;
It contains axons and dendrites of the cells of the underlying layers, as well as stellate and basket cells (Fig. 32,
1
).
2) middle - ganglion layer
(
stratum ganglionaris
)
;
Formed by large pear-shaped Purkinje cells, which have a powerful, highly branching dendritic tree in the molecular layer (Fig. 32, 5
).
3) internal - granular layer
(
stratum granulosum
).
Axons of granule cells are directed to the molecular layer, where they branch in a T-shape and enter into synaptic contacts with the dendrites of Purkinje cells, basket and stellate cells (Fig. 32, 4
).
The dendritic tree of Purkinje cells is located in a plane perpendicular to the axis of the furrow, and the axons of granule cells are parallel to it. There are about 5 thousand granule cells per Purkinje cell. On the soma and dendrites of Purkinje cells, the axons of stellate and basket cells, as well as the so-called climbing fibers from the olive nuclei (which came to the cerebellar cortex along the olivo-cerebellar tracts) also end. The remaining afferent pathways end in the cerebellar cortex in the form of mossy fibers (Fig. 32, 5) on granule cells, as well as on stellate and basket cells. Efferent outputs from the cerebellar cortex are created by axons of Purkinje cells ending on cells of the subcortical nuclei of the cerebellum. The axons of the cells of the cerebellar nuclei make up the efferents of the cerebellum, connecting it with other parts of the central nervous system.
Afferent and efferent fibers together form three pairs of cerebellar peduncles. Through the lower pair of peduncles, the cerebellum receives afferents from the dorsal spinocerebellar tract of Flexing, here pass the olivo-cerebellar tract, pathways from the vestibular nuclei of the VIII pair of nerves and the nuclei of the V, VII, IX and X pairs of cranial nerves, as well as from the Gaulle and Burdach nuclei of the medulla oblongata . Through the lower legs, only one efferent pathway emerges from the nuclei of the tent to the vestibular nuclei of the medulla oblongata. The middle legs have only afferent fibers coming from the own nuclei of the bridge, as well as collaterals from the corticospinal tracts. Through these legs, various sections of the cerebral cortex (frontal, temporal and occipital) are connected to the cerebellum, since the corticopontine descending pathways end on the pons' own nuclei. Through the superior peduncles, the cerebellum receives afferent fibers from the ventral spinocerebellar Gowers tract, as well as from the anterior colliculus. The bulk of the anterior legs consists of efferent fibers going to the red nucleus, reticular nuclei and colliculus of the midbrain, to the thalamic and hypothalamic nuclei of the diencephalon. Through the thalamic nuclei, the cerebellum is connected to the cerebral cortex, and through the red nuclei, nuclei of the reticular formation and vestibular nuclei - to the spinal cord.
Phylogenesis.
Phylogenesis.
Among vertebrates, the most primitive is the cerebellum of cyclostomes. It is a plate with an outer layer of fibers and an inner cell layer, which is connected to the lateral line organ and the vestibular nuclei. Subsequently, the development of the cerebellum was determined by the improvement of vestibulocerebellar connections. Therefore, in fish, the cerebellum develops in such a way that it becomes the highest integrative structure. The cerebellum of fish consists of a body and two small elevations. Cyclostomes already have elevations. They are called the ancient cerebellum, and the body is called the old cerebellum. When animals reach land, the cerebellum is first reduced (in amphibians), and then again develops into a powerful brain structure (in reptiles and birds). Development follows the path of improving connections between the spinal cord and the cerebellum and weakening connections with the vestibular system. In higher reptiles (crocodiles) and birds, the cerebellar cortex is formed, which has two layers of cells: granular and molecular (with Purkinje cells). The body of the cerebellum is divided into three lobes: anterior, middle and posterior. In mammals, structures arise in the cerebellum that are closely connected with the cerebral cortex - the new cerebellum. The cerebellar hemispheres, as well as its middle peduncles, appear for the first time. They are formed from the body of the cerebellum of lower vertebrates. In rodents, three subcortical nuclei appear in the cerebellum (tent, dentate and median). And only in primates the median nucleus splits into spherical and cork-shaped. The dentate nucleus reaches its greatest development.
The developed cerebellum of higher mammals consists of three parts: ancient ( paleocerebellum
) controls vestibular function (its caudal sections);
the old one ( archicerebellum
) is connected to the spinal cord (anterior lobe) and the new one (
neocerebellum
) is connected to the cerebral cortex.
Ontogenesis.
Ontogenesis.
In ontogenesis, the cerebellum develops from the cerebellar plate, which is the roof of the metencephalic vesicle (
metencephaton
). The cerebellar hemispheres are formed from the lateral parts, and the vermis is formed from the middle part of this plate.
Return to contents
© 2000-NIV
Functions of the hindbrain area
The responsible area of the nervous system, in which, thanks to the action of the nuclei, the chains of a large number of vegetative and somatic reflexes are closed, is the hindbrain. These include, for example, chewing and swallowing reflexes; regulation of the intensity of salivation as a secretion of the salivary glands.
The hindbrain, namely the suprasegmental organ, the cerebellum, is responsible for such specific actions as regulating the tone of different muscle groups; sensorimotor coordination of body position and meaningful movements; implementation of instant, purposeful movements based on impulses from the cerebral cortex of the cerebral hemispheres. When any disturbances in the functioning of the hindbrain appear, certain pathological symptoms arise: unnecessary movements, alternating paralysis, legs unnaturally wide apart when walking, swaying from side to side, etc.
Functions of the pons
The hindbrain, the functions of the pons in which support the activity of the entire organism, primarily controls muscle contractions and stability when walking. The bridge contains nerve fibers that transmit impulses to the cortex and from the cerebral cortex in the opposite direction - to the spinal cord, medulla oblongata, and cerebellum. There are the main centers that control facial expressions, chewing functions of a person, hearing and vision. The pons is even responsible for some reflexes: blinking, coughing, sneezing, vomiting. Therefore, we can say that the only purpose of the bridge is to transmit the necessary information from the spinal cord to the large brain, and then to some internal organs.
What is the midbrain responsible for?
Memory is a complex process that includes three phases: encoding (deciding what information is important), storage, and recall. Different areas of the brain are involved in different types of memory
Your brain must pay attention and rehearse for the event to move from short-term memory to long-term memory—called encoding. Fig 6
The structures of the limbic system are involved in memory formation. The prefrontal cortex briefly holds recent events in short-term memory. The hippocampus is responsible for encoding long-term memory.
Short-term memory, also called working memory, originates in the prefrontal cortex of the brain. It stores information for approximately one minute and its capacity is limited to approximately 7 items. For example, it allows you to dial a phone number that someone just told you. It also intervenes during reading to remember the sentence you just read so that the next one makes sense.
The midbrain is located in the anterior pons and extends to the papillary bodies, as well as to the optic tracts. Here, clusters of nuclei are identified, which are called quadrigeminal tubercles. The structure and functions of the brain sections (table) indicate that this section is responsible for latent vision, the orientation reflex, gives orientation to reflexes to visual and sound stimuli, and also maintains the tone of the muscles of the human body.
The medulla oblongata is a natural extension of the spinal cord. That is why there are many similarities in the structure. This becomes especially clear if we examine the white matter in detail. Its short and long nerve fibers represent it. Gray matter is represented here in the form of nuclei. The parts of the brain and their functions (the table above) indicates that the medulla oblongata controls our balance, coordination, regulates metabolism, controls breathing and blood circulation. It is also responsible for such important reflexes of our body as sneezing and coughing, vomiting.
Only parts of the organ’s cortex, the limbic system and the cerebellum are responsible for memory functioning. The main influence is on areas located in the temporal zone of the left and right hemispheres.
Also, the main department for storing long-term information is the hippocampus.
He is responsible for cross-functional activities. Conveys motor sensations (coordination), tactile, and reflex sensations.
With the help of this area, a person can move in space without any problems.
The left hemisphere, in which the speech zones are located - motor and sensory - is mainly responsible for speech function.
hindbrain
The pons, which from an anatomical point of view is considered a structure of the hindbrain, is presented in the form of a thickened cushion. At the bottom of the bridge there is an oblong section, at the top there is a middle section.
The pons contains centers that control the functioning of the masticatory, facial, and some oculomotor muscles. Nerve impulses from the receptors of the sensory organs, skin, and inner ear go to the bridge; thanks to this zone, we can feel taste, maintain balance, and have hearing sensitivity.
Structure and functions of the cerebellum
The lower part of the cerebellum is adjacent to the medulla oblongata, which is a continuation of the spinal cord and consists of two brain substances - gray and white. Both hemispheres of the cerebellum are separated from the cerebrum by a deep horizontal fissure, and the surface is dotted with fissures and convolutions of the medulla, forming the anterior, posterior and flocnonodular lobes. The hemispheres are connected to each other by a worm, and inside them lie the nerve nuclei.
The cerebellar gray matter seems to branch out into the white, resembling a twig of thuja. Located at the edges, the gray forms a layered cortex, under which there is always white medulla. Coordination of movement and balance during movement depend on the proper functioning of the cerebellum, and the main function of the department is to connect the forebrain with the hindbrain.
Cerebellum
The cerebellum is a structure located at the back of the brain responsible for sensorimotor functions. Located behind the cerebral hemispheres of the telencephalon above the dorsal surface of the medulla oblongata and the pons. Anatomically, the cerebellum is a separate structure of the brain, visually standing out against its background. It is the main structure of the hindbrain. The cerebellum is connected to the pons by the anterior cerebellar peduncles, which are wide bundles of nerve fibers.
The surface of the cerebellum is penetrated by thin parallel grooves, which are noticeably different from the large convolutions of the cerebral hemispheres. These grooves are a thin continuous layer of the cerebellar cortex, as if folded several times, like an accordion. It is believed that if you unfold the cerebellar cortex and spread out this nervous tissue, smoothing out all the folds, it would occupy an area of approximately 500 square centimeters. This layer contains several types of densely packed neurons, the most important of which are Purkinje cells and granule cells. This complex neural structure facilitates the processing of a large array of signals. Most outputs from the cerebellar cortex pass through the small nuclei, which are located in the white internal substance of the cerebellum.
It should be noted that the cerebellum consists of two hemispheres, which are connected through a “worm” - an unpaired structure. The cerebellum occupies most of the posterior cranial fossa. The average weight of the cerebellum in an adult is about 140 grams. The cerebellum contains a large number of neurons, comparable to the number of neurons in the cerebral cortex.
The cerebellum is connected to various parts of the nervous system through three cerebellar peduncles: superior, inferior and middle. The legs are named after their location relative to the worm. The superior cerebellar peduncle extends to the cerebral cortex, carrying efferent fibers arising from the deep cerebellar nuclei to the upper motor neurons in the cerebral cortex. The middle cerebellar peduncle is connected to the pons, receiving all incoming information from it, which mainly comes from the Pontini nuclei. This pedicle is the largest bundle of cerebellar fibers. The inferior cerebellar peduncle receives input from afferent fibers from the vestibular nuclei, spinal cord, and tegmentum. The exit from the inferior peduncle occurs through efferent fibers in the vestibular nuclei and reticular formation. The entire cerebellum also receives modulating input from the inferior olivary nucleus via the inferior cerebellar peduncle.
Functions and significance of the medulla oblongata
The hindbrain, whose functions are so important for the functioning of the body, is in close contact with the medulla oblongata, which is responsible for many processes. Nerve impulses that pass from the cerebrum to the spinal cord and then in the opposite direction are transmitted through the pons and medulla oblongata.
The peculiarity of the structure of the latter is that it reads information from sensory organs and nerve impulses and is responsible for metabolism in the body. The centers for regulating the digestive organs and some others are located here.
In addition, the medulla oblongata plays a critical role in controlling breathing and sweating, as well as the state of the cardiovascular system. It is in this department that the responsible centers are located, excited not only reflexively by receiving nerve impulses from the periphery, but also by means of internal chemical stimuli acting on them.
The forebrain and hindbrain are in close interaction, fully responsible for the functioning of the body as a whole. It is thanks to brain cells (neurons) that impulses are transferred, and knowledge of the paths in the hindbrain helps to understand the mechanisms of the appearance of functional pathologies and disorders and, if possible, eliminate them.
Hindbrain (Metencephalon)
Bridge
The bridge (pons) is an elevation located between the medulla oblongata and midbrain, 25-27 mm long. Its lower border is the pyramids and olives of the medulla oblongata, the upper is the cerebral peduncle, the lateral is the line passing between the roots of the trigeminal and facial nerves. On the dorsal side, the upper border of the bridge is the upper cerebellar peduncles (redunculi cerebellares superiores) and the superior medullary velum (velum medulla-re superius), and below there is a deep horizontal groove from which, starting from the main groove, the roots of the abducens (VI pair) emerge. facial (VII pair) and auditory (VIII pair) nerves.
The bridge is divided into front and rear parts. The anterior part of the bridge (pars anterior pontis) is convex and formed by transverse nerve fibers connecting the cells of the cerebral cortex with the nuclei of the bridge (nucll. pontis) and then with the cerebellar cortex. Together with them, fibers go in the opposite direction from the cerebellar cortex to the cerebral cortex. These fibers cover the bundles of the pyramidal tract running perpendicularly (Fig. 465), and then in the lateral parts of the pons they are collected into the middle cerebellar peduncles (pedunculi cerebellares medii). Along the midline of the bridge between the elevations formed by the fibers of the pyramidal tract, there is a basilar groove (sulcus basilaris), in which the artery of the same name lies.
The dorsal part of the bridge is thinner and participates in the formation of the upper part of the rhomboid fossa. In the dorsal part of the bridge are located the nuclei of the V, VI, VII, VIII cranial nerves, the reticular formation, and the superior olive. The latter is connected with the auditory nuclei and has connections with the reticular formation of the medulla oblongata and midbrain.
The sensory and motor nuclei of the trigeminal nerve (V nary) are located in the upper part of the bridge. The sensitive nucleus (nucleus sensorius n. trigemini) is the site of switching of cell processes of the trigeminal ganglion. The motor nucleus (nucl. motorius n. trigemini) consists of small pyramidal cells that innervate the masticatory muscles.
The nucleus of the abducens nerve (nucl. n. abducentis) (VI pairs) is located in the lower part of the bridge near the midline.
The nucleus of the facial nerve (nucl. n. facialis) is formed by motor cells that innervate facial muscles. They are located in a reticular formation. The fibers of the nucleus form a genu that bends around the nucleus of the abducens nerve. Behind the motor nucleus of the facial nerve lies the superior salivary nucleus (nucl. salivatorius superior), where the fibers for the innervation of the lacrimal, sublingual and submandibular glands begin. Lateral to the superior salivary nucleus is the nucleus of the solitary tract (nucl. tr. solitarii), (nucleus of the VII pair), which has the shape of a column reaching the medulla oblongata. Sensitive fibers of the cells of the ganglion geniculi, which are conductors of taste sensations, end in the nucleus.
The nuclei of the vestibular-cochlear nerve (n. vestibulocochlearis) are located in the inferolateral part of the posterior part of the bridge.
Olives. The superior olive (oliva superior) has nuclei lying in the lateral sections of the bridge at the level of the trapezoid body, i.e., on the border of its ventral and dorsal parts.
The reticular formation (formatio reticularis) has several nuclei, predominantly oriented in the cross-sectional plane (Fig. 465).
465. Diagram of the location of conductive paths and nuclei on a cross section of the bridge. 1 - nuclei of the V pair; 2 - nuclei of the VIII pair; 3 - tr. rubrospinal; 4 - tr. spinocerebellaris anterior; 5 - tr. spinocerebellaris posterior; 6 - tr. spinothalamic lateralis; 7 -VII pair; 8 - VI pair; 9 - tr. corticospinalis (pyramidalis); 10 - fasc. longitudinalis medialis; 11 - tr. spinothalamicus anterior; 12 - tr. tectospinal; 13 - tr. reticulospinalis
1. The lateral reticular nucleus (nucl. reticularis lateralis) lies lateral and below the inferior olive. Sends its fibers through the opposite inferior cerebellar peduncles to the cerebellum.
2. The reticular nucleus of the tegmentum of the bridge (Bechterew's spondylitis) (nucl. reticularis tegmenti pontis) surrounds the own core of the bridge. Some of its fibers reach the cerebellar vermis, others, crossing, end in the cerebellar hemispheres.
3. The paramedial reticular nucleus (nucl. paramedialis) is located medial and dorsal to the inferior olive. Some of the fibers cross and reach the vermis, hemispheres and the tent nucleus of the cerebellum.
4. The reticular giant cell nucleus (nucl. retucularis gigantocel-lularis) represents 2/3 of the volume of the reticular formation. It is located dorsal to the superior olive, extending upward to the nucleus of the facial nerve. Long processes of the cells of the giant cell nucleus reach the overlying and underlying parts of the brain.
5. The caudal reticular nucleus (nucl. reticularis caudalis) is located above the previous one.
6. The oral reticular nucleus (nucl. reticularis oralis) is located on the border with the midbrain. Continues into the mesencephalic reticular formation. The fibers of the caudal and oral nuclei, together with the fibers of the giant cell nucleus, form ascending and descending fiber systems.
The trapezoidal body (corpus trapezoideum) is located between the front and rear parts of the bridge in the form of a strip 2-3 mm wide. Formed by the own nuclei of the trapezoidal body (nucl. proprius), as well as fibers of the ventral and dorsal auditory nuclei (nucl. cochleares anterior et posterior). The processes of the cells of the nuclei of the trapezoid body, the anterior and posterior nuclei are combined into a lateral loop (lemniscus lateralis), which also has its own nucleus (nucl. lemnisci lateralis). The trapezoid body, the anterior and posterior nuclei, and the lateral lemniscus participate in the formation of the auditory pathway.
Age characteristics. The bridge in newborns lies 5 mm above the back of the sella turcica. By 2-3 years it descends to the slope of the skull. The nuclei of the cranial nerves are well differentiated, the fibers of the corticospinal tracts become covered with myelin by the age of 8 years.