Добрый день, уважаемые читатели. Сегодня мы изучим проводящие пути среднего мозга. Перед началом работы с нашей сегодняшней темой, обязательно прочитайте мои руководства по спинному мозгу, продолговатому мозгу и мосту.
Если вы будете соблюдать эту очередность и изучать части ЦНС по моим статьям, вам не придётся потом отдельно учить проводящие пути — вы их и так будете знать, потому что в каждом новом уроке я провожу связи со структурами из предыдущих уроков.
Что такое проводящий путь? Проводящий путь — это последовательность нейронов, соединённых между собой для проведения импульса в строго определённом направлении. Например, путь Говерса, который обеспечивает проведение неосознанной проприоцептивной чувствительности, состоит из двух нейронов:
- Тело первого нейрона находится в спинномозговом ганглии, а аксон уходит в мышцы / суставные капсулы / сухожилия;
- Тело второго нейрона находится в заднем роге спинного мозга, аксоны его уходят в белое вещество спинного мозга; затем проходя через продолговатый мозг и мост, заходят в мозжечок;
- Тело третьего нейрона находится в коре червя мозжечка.
Когда мы говорим о теле и отростке какого-либо нейрона, мы понимаем, что в действительности речь идёт не об одном, а об огромном количестве тел нейронов (их скопление называется ядром) и целых пучках отростков нейронов. На уровне каждой части ЦНС имеются свои ядра и свои пучки волокон. Например, в прошлом уроке мы уже изучили ядра среднего мозга. Теперь мы будем изучать расположение пучков волокон, проходящих через средний мозг. В анатомических учебниках пучки волокон, формирующих проводящие пути, называют путями, трактами, или же просто проводящими путями.
Проводящие пути среднего мозга
Как и проводящие пути других нижележащих участок ЦНС, пути среднего мозга делят на:
- Чувствительные (проводят информацию от рецепторов, например, палочек/колбочек, к центральной нервной системе);
- Двигательные (проводят информацию к исполнительным органам, лежащим вне ЦНС, например, к скелетным мышцам);
- Ассоциативные (соединяют участки одной и той же части ЦНС между собой).
Однако, в среднем мозге есть ещё одна группа необычных проводящих путей, которая связана с наличием черного вещества. Черное вещество, как вы помните из прошлого гайда, синтезирует дофамин и передаёт его в вышележащие структуры головного мозга. Мы обязательно разберём и эти проводящие пути, но начнём с более привычных и уже известных нам двигательных и чувствительных путей среднего мозга.
Двигательные пути среднего мозга
Красноядерно-спинномозговой тракт
Когда вы смотрите на разрез среднего мозга, вы не можете не заметить крупные, контрастные структуры, называемые красными ядрами. Красные ядра являются одним из ключевых компонентов экстрапирамидной двигательной системы — то есть системы неосознанных движений, которые не требуют тщательно обработки в коре головного мозга.
От красных ядер вниз, к спинному мозгу и, через него — к скелетным мышцами, идёт путь, называемый красноядерно-спинномозговым путём (tractus rubrospinalis). Также существуют названия:
- Монаковский пучок;
- Руброспинальный тракт —
— это равноценные названия одного и тоже же проводящего пути.
Обязательно отмечаем перекрест волокон руброспинального, который располагается здесь же, на уровне среднего мозга. Точка пересечения двух руброспинальных путей имеет своё название — передний покрышечный перекреcт (decussatio tegmenti anterior). По автору это анатомические образование называется перекрестом Фореля.
Это похоже на лесенку, потому что в прошлых уроках я обозначал руброспинальный путь именно так. Я стараюсь выдерживать одни и те же проводящие пути в одной цветовой гамме, чтобы нам было проще их собирать.
Крыше-спинномозговой тракт
Это ещё один путь экстрапирамидной системы. Крыше-спинномозговой путь (tractus tectospinalis) начинается от чувствительных ядер, залегающих в верхних и нижних холмиках четверохолмия. Как вы помните, там залегают, соответственно, подкорковые центры зрения и слуха.
В нашем случае термин «подкорковые» означает, будет происходить быстрая, моментальная обработка зрительных и слуховых сигналов. После этой обработки будет произведено переключение на двигательные нейроны крыше-спинномозгового пути.
Представьте, что в комнате, где вы сейчас находитесь, упадёт металлический поднос вне вашего поля зрения. Вы неожиданно услышите внезапный, резкий грохот. Первой реакцией вашего организма будет непроизвольное сокращение скелетных мышц — вы вздрогнете или даже подскочите. Именно эту реакцию и обеспечивает крыше-спинномозговой тракт.
Похожая ситуация и со зрением — если в поле вашего зрения мелькнёт какой-то объект, вы невольно развернёте голову в его сторону. Если движение объекта будет слишком близком к вам, вы, скорее всего, на долю секунды зажмурите глаза и закроете лицо руками.
Именно эти реакции и являются подкорковыми. Давайте нарисуем наш тектоспинальный путь, не забыв дорисовать первые нейроны. Конечно, я сильно ошибся, не нарисовав их в прошлом уроке про ядра среднего мозга:
Как вы видите, крыше-спинномозговой тракт также образует перекрест. Этот участок пересечения называется задним покрышечным перекрестом (decussatio tegmenti posterior), а по автору — перекрестом Мейнерта.
Медиальный продольный пучок
Отростки тел нейронов, формирующих ядра Кахаля и Даршкевича, соединяются в пучки, которые называются медиальными продольными пучками (fasciculus longitudinalis medialis). Как вы помните, этот проводящий путь анастомозирует с ядрами черепных нервов, отвечающих за движение глаз, а также с двигательными волокнами спинного мозга. В итоге, медиальный продольный пучок отвечает за сочетанные движения головы и глаз.
Нарушение этой функции очень схоже с тем, как выглядит списывающий студент, который развернул голову в сторону преподавателя, а глаза — в сторону своих колен и гаджета со шпаргалками.
Лобно-мостовой путь
Сейчас мы переместимся в ножки мозга, которые, по сути, представляют собой выступающие, оформленные участки белого вещества. Медиальнее всех здесь будут располагаться волокна лобно-мостового пути (tractus frontopontinus). Как вы помните, в базилярной части моста мы наблюдали множество мелких, рассеянных по значительной площади ядер, которые назывались собственными ядрами моста. Аксоны нейронов, которые группировались в эти ядра, формировали средние мозжечковые ножки и направлялись, соответственно, в мозжечок.
Оказывается, эти аксоны и ядра получают информацию от нейронов коры головного мозга. Совокупность волокон, которые соединяют кору лобной доли и собственные ядра моста, называют лобно-мостовым путём.
В отличие от ранее изученных нами трактов, лобно-мостовые пути не начинаются от ядер среднего мозга, а проходят через него транзитом. Эти пути располагаются медиальнее всех остальных трактов ножек мозга:
Затылочно-висично-мостовой путь
Затылочно-висично-мостовой путь(tractus occipitotemporopontinus) путь полностью аналогичен лобно-мостовому пути, за тем исключением, что в этом случае тела первых нейронов находятся не в лобной извилине, а височной и затылочной извилинах. Кроме того, пучки этого пути проходят не в медиальной, а в наиболее латеральной части ножек мозга. На этом отличия заканчиваются:
Корково-спинномозговой путь
Вот, наконец, мы и добрались до кортико-спинальных путей (tractus corticospinalis), которые в нижележащих структурах переходят во всем известные пирамиды.
Корково-спинномозговые пути начинаются из клеток коры головного мозга. Эти клетки залегают в 5-м слое коры и называются пирамидными клетками Беца. Прилагательное «пирамидный» здесь употребляется из-за некоторого сходства этих нейронов с пирамидами. Это хорошо заметно при рассматривании микроскопических изображений пирамидных клеток Беца.
Корково-спинномозговые пути являются очень важным эволюционным приспособлением. Именно пирамидная система (иногда проводящие пути, начинающиеся от пирамидных клеток, называют пирамидной системой) позволяет нам совершать осмысленные движения с предварительным глубоким, качественным корковым анализом. Например, когда я впервые попробовал завязать хирургический узел (это был узел по Джанелидзе), мне приходилось очень долго думать перед каждым движением. Вот тут-то моя пирамидная система работала в полную мощность!
Зато потом, после нескольких тысяч повторений, завязывание этого узла перестало требовать такого долгого и тщательного анализа со стороны пирамидной системы и в дело частично включилась экстрапирамидная система.
Волокна кортикоспинального пути, начинаясь от клеток Беца, спускаются вниз, проходя транзитом через нижележащие структуры головного и спинного мозга. Как вы помните, на уровне продолговатого мозга эти тракты совершают перекрест, после которого 80% волокон оказываются на противоположной стороне. Но на уровне среднего мозга никаких перекрестов нет, посему мы просто закрашиваем часть ножки мозга, в которой залегает корково-спинномозговой путь:
Примечание 1: наличие поперечных полосок на моём рисунке — это просто необходимость как-то визуально отделять пути ножек мозга друг от друга при том, что, согласно анатомической традиции, все двигательные пути должны быть красными, а все чувствительные — синими.
Примечание 2: топографически мой рисунок не совсем правильный, потому что кортикоспинальные тракты должны занимать самое большое пространство ножки мозга из всех остальных путей.
Корково-ядерный путь
Корково-ядерный путь (tractus corticonuclearis) — это ещё один тракт, формирующий пирамидную систему. Однако, в отличие от корково-спинномозгового пути, инневирующего скелетные мышцы туловища и конечностей, корково-ядерный путь иннервирует скелетные мышцы лица — жевательные и мимические, а также мышцы шеи.
Чувствительные пути среднего мозга
Наш средний мозг выглядит слишком красным, не так ли? Немного голубого — традиционного цвета чувствительных проводящих путей — здесь явно не помешает.
Спинно-таламический путь
Хорошо знакомый нам ещё с предыдущих уроков, спинно-таламический путь (tractus spinothalamicus) отвечает за проведение тактильных, температурных и болевых ощущений с кожных рецепторов. На уровне мезенцефалона спинно-таламический путь не переключается ни на какие ядра, не начинается и не заканчивается, поэтому мы отмечаем пучок волокон в виде горизонтального среза:
Медиальная петля
Медиальная петля (lemniscus medialis) — это волокна проводящих путей осознанной проприоцептивной чувствительности. На уровне спинного мозга эти пути называются путями Голя и Бурдаха. Далее, совершая перекрест на уровне миеленцефалона, эти пути соединяются в один тракт, называемый медиальной петлёй. В виде медиальной петли эти волокна будут продолжать подниматься до таламуса, где переключатся на нейроны, направляющиеся в кору больших полушарий.
На уровне среднего мозга никаких переключений не происходит, мы видим просто пучки волокон:
Латеральная петля
Латеральная петля (lemniscus lateralis) — это волокна проводящего пути, который связывает орган слуха и участки центральной нервной системы. На уровне среднего мозга часть волокон этого пути располагается в подкорковых центрах, то есть, нижних холмиках пластинки четверохолмия, а остальная часть направляется прямо в большие полушария, точнее — в кору верхней височной извилины.
В действительности, латеральная петля не настольно широка относительно медиальной петли, просто мне пришлось её немного «расплющить» из-за недостатка пространства внутри рисунка. В атласах латеральную петлю изображают в виде
Пути транспортировки дофамина. Нигростриарный путь
В мезенцефалоне располагается анатомическое образование, называемое чёрным веществом. Нейроны черного вещества синтезируют дофамин — гормон и нейромедиатор, который осуществляет передачу импульса в путях, отвечающих, в том числе, за возникновение чувства удоволетворения от работы и мотивации к действию. Также дофамин является одним из медиаторов экстрапирамидной системы.
С нарушением синтеза дофамина в черном веществе несомненно связано грозное неврологическое страдание — болезнь Паркинсона. Также с метаболизмом дофамина, предположительно, связана ещё одна инвалидизирующая болезнь — шизофрения.
Для того, чтобы доставлять дофамин в вышележащие структуры больших полушарий, нейроны черной субстанции отправляют свои отростки к полосатому телу — мы обязательно разберём это анатомическое образование позже. А сейчас мы должны запомнить, что значительная часть отростков нейронов чёрного вещества направляется наверх, к большим полушариям и несёт туда дофамин. За это отвечают три пути, самым главным из которых является нигростриарный путь (tractus nigrostriatalis). Я схематично отметил ход нигростриарного пути на нашей иллюстарции:
Текст книги «Клиническая диагностика в неврологии»
Глава 2 РАССТРОЙСТВА НЕПРОИЗВОЛЬНЫХ И ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
2.1. Виды движений
Движения подразделяются на непроизвольные и произвольные. Непроизвольные движения
– это движения, возникающие независимо от желания человека в ответ на воздействие различных раздражителей. Подобные рефлекторные движения присущи каждому биологическому виду, формируются к моменту рождения, передаются по наследству. Их анатомо-физиологическим субстратом является рефлекторная дуга или рефлекторное кольцо, замыкающиеся на уровне спинного мозга или ствола головного мозга.
Произвольные движения
представляют собой движения различной степени сложности, подразделяющиеся на три группы:
1. Элементарные сознательно-волевые движения.
Представляют собой дифференцированные двигательные акты, которые являются произвольными и при этом имеют относительно простой характер – поднятие руки или ноги, их сгибание в суставах и т. д. Эти движения «запускаются» на основе эфферентного импульса, формирующегося в первичном корковом поле двигательного анализатора (преимущественно прецентральная извилина).
2. Двигательные праксисы.
Это более сложные двигательные акты, которые вырабатываются в процессе жизни человека на основе опыта, навыка, практики и закрепляются стереотипом. Программирование подобных движений (причесывание, застилание постели, письмо и т. д.) осуществляется в особых зонах коры (вторичные, или ассоциативные), а их реализация осуществляется также через первичное корковое поле двигательного анализатора и его эфферентные связи.
3. Автоматизированные движения
– бег, ходьба, ползание, плавание и т. д. – произвольны относительно, поскольку их выполнение имеет заученный характер и осуществляется как единый двигательный акт. Подобные движения являются прерогативой преимущественно экстрапирамидной системы и мозжечка, а двигательные поля коры головного мозга оказывают на них в основном регулирующее действие.
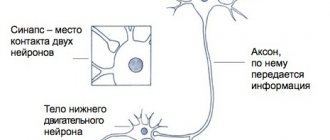
2.2. Центральный и периферический двигательные нейроны корково-мышечного пути
Для осуществления произвольного движения необходимо, чтобы двигательный импульс, возникший в соответствующих областях коры головного мозга, был проведен к скелетным (поперечно-полосатым) мышцам. Это обеспечивает корково-мышечный путь (tractus corticomuscularis)
, состоящий из двух частей: первая часть представлена центральным двигательным нейроном; вторая – периферическим двигательным нейроном.
Центральный двигательный нейрон.
Согласно классическим представлениям тела нейронов, от которых берет начало корково-мышечный путь, располагаются в передней центральной извилине – первичном корковом поле двигательного анализатора. Долгое время считалось, что центральный двигательный нейрон начинается только от тел внутренних больших пирамидных нейронов (клеток Беца), расположенных в пятом слое передней центральной извилины, что и предопределило его название – пирамидный путь. Согласно данным последних лет, от передней центральной извилины начинается только 27 – 40 % аксонов корково-мышечного пути, а непосредственно от клеток Беца – лишь 3 – 4 %, около 20 % волокон корково-мышечного пути берут начало от соматосенсорной корковой зоны (постцентральная извилина), а остальные – от премоторной зоны, парацентральной и других областей мозга. Аксоны этих нейронов заканчиваются на мотонейронах передних рогов спинного мозга (корково-спинномозговой путь —
tractus corticospinalis
) и на двигательных ядрах черепных нервов (корково-ядерный путь —
tractus corticonuclearis
).
Периферический двигательный нейрон
образован мотонейронами передних рогов спинного мозга и двигательных ядер черепных нервов, а также их аксонами, достигающими в составе ряда образований нервной системы (передние корешки, спинномозговые нервы, сплетения, периферические или черепные нервы) мышц-исполнителей.
2.3. Анатомия корково-спинномозгового и корково-ядерного путей
Корково-спинномозговой путь
начинается преимущественно от верхних 2/3 передней центральной извилины и парацентральной дольки; часть аксонов берет начало в прилегающей премоторной области, в задней центральной извилине и верхней теменной дольке (рис. 2.1).
Рис. 2.1.
Схема пирамидных путей:
1 – латеральный (перекрещенный) пирамидный пучок; 2 – прямой (неперекрещенный) пирамидный пучок; 3 – корково-ядерные волокна; 4 – проекция двигательной сферы в передней центральной извилине коры головного мозга
В коре передней центральной извилины пирамидные нейроны локализуются по правилу «моторного гомункулюса» (схема Пенфилда). Это означает, что в самых верхних отделах передней центральной извилины располагаются нейроны, которыми начинаются эфферентные пути для мышц ноги: в парацентральной дольке располагаются нейроны, обеспечивающие движения мышц стопы, а в верхних отделах передней центральной извилины – последовательно соматотопическая проекция для мышц голени и бедра. Далее последовательно располагаются нейроны, дающие начало эфферентным нервным путям к мускулатуре туловища. Среднюю треть передней центральной извилины занимают нейроны, обеспечивающие иннервацию мышц руки. Площадь соматотопических проекционных зон в передней центральной коре пропорциональна сложности движений, выполняемых определенной группой мышц, причем наибольшую площадь занимает соматотопическая проекция мышц кисти (особенно мышц тенара).
Аксоны из вышеуказанных областей передней центральной извилины проходят через лучистый венец, передние 2/3 заднего бедра внутренней капсулы и далее следуют по вентральной поверхности ствола головного мозга. На границе между продолговатым и спинным мозгом 80 % волокон корково-спинномозгового пути переходит на противоположную сторону, образуя с аналогичными волокнами противоположной стороны пирамидный перекрест (decussatio pyramidum)
. Перекрещенные волокна направляются в боковой канатик спинного мозга, занимая его заднемедиальный отдел в виде латерального корково-спинномозгового пути
(tractus corticospinalis lateralis)
. Этот путь обеспечивает произвольные движения как мышц туловища, так и конечностей. Около 20 % волокон остаются неперекрещенными, образуя передний корково-спинномозговой путь
(tractus corticospinalis anterior)
. Этот путь обеспечивает произвольные движения преимущественно в мышцах туловища и шеи. Волокна латерального корково-спинномозгового пути располагаются в спинном мозге в соответствии с законом Ауэрбаха – Флатау и при этом посегментно переключаются на мотонейроны передних рогов спинного мозга.
Корково-ядерный путь
начинается преимущественно от нижней трети передней центральной извилины и прилегающей премоторной области. Здесь также определяется четкая соматотопическая проекция мышц, иннервируемых черепными нервами, причем в самом нижнем отделе прецентральной извилины располагаются нейроны для мышц глотки, гортани, мягкого нёба, языка, жевательной и мимической мускулатуры. Аксоны корково-ядерного пути проходят через лучистый венец, колено внутренней капсулы и ствол мозга. Здесь его волокна совершают частичный надъядерный перекрест, заканчиваясь на двигательных ядрах черепных нервов своей и противоположной стороны. Исключение составляет часть корково-ядерного пути, заканчивающаяся на ядре XII и нижней части ядра VII нерва – эти волокна совершают полный надъядерный перекрест и поэтому заканчиваются на соответствующих двигательных ядрах (ХII, нижняя половина VII) только противоположной стороны.
2.4. Плегия и парез
Плегия, или паралич,
– полная утрата произвольных движений в той или иной группе мышц.
Парез
– частичная утрата произвольных движений в той или иной группе мышц, характеризующаяся снижением силы и объема активных движений в пораженных мышцах.
В зависимости от распространенности различают следующие варианты параличей и парезов:
– моноплегия, или монопарез, – плегия или парез определяются только в мышцах одной руки или ноги;
– гемиплегия, или гемипарез, – плегия или парез захватывает мышцы как руки, так и ноги на одной или на противоположных (альтернирующая гемиплегия или гемипарез) сторонах тела;
– параплегия, или парапарез, – плегия или парез определяются только в мышцах только обеих рук или ног (соответственно верхняя или нижняя параплегия или парапарез);
– триплегия, или трипарез, – плегия или парез захватывает три конечности;
– тетраплегия, или тетрапарез, – плегия или парез захватывает обе руки и ноги.
2.5. Общие клинические признаки расстройств движений
Плегии (парезы) могут быть выявлены при клиническом осмотре на основании следующих признаков:
1) снижение объема активных движений и (или) мышечной силы;
2) изменение мышечного тонуса;
3) атрофия или гипотрофия паретичных мышц;
4) фасцикуляции и фибрилляции пораженных мышц;
5) повышение или снижение (вплоть до отсутствия) физиологических рефлексов, замыкающихся на паретичных мышцах;
6) наличие патологических рефлексов;
7) наличие защитных рефлексов и патологических синкинезий. В зависимости от вида плегии или пареза – периферический или центральный – различные вышеперечисленные симптомы образуют клиническое ядро периферического или центрального паралича.
2.6. Методика исследования силы мышц различных мышечных групп
Мышечная сила исследуется параллельно с активными движениями, так как их объем при нерезко выраженных парезах не уменьшается. Силу мышц кистей определяют динамометром. При исследовании силы других мышц используют ручной способ в двух модификациях.
При первой модификации врач оказывает препятствие в выполнении активного движения, определяет и сравнивает слева и справа силу сопротивления в соответствующих мышцах. Так, например, врач предлагает больному сгибать руку в локтевом суставе слева и активно препятствует этому сгибанию. Затем так же определяется сила двуглавой мышцы плеча правой руки, и сила активного движения слева и справа сравнивается.
Чаще используют другую модификацию: пациенту предлагают выполнить активное движение, не оказывая противодействия. Далее пациент удерживает руку или ногу в этой позе с максимальной силой, а врач пытается произвести движение в обратном направлении. При этом он оценивает и сравнивает слева и справа степень усилия, которое для этого требуется. Например, силу двуглавой мышцы плеча определяют, пытаясь разогнуть уже согнутую в локтевом суставе руку, вначале слева, затем справа.
2.7. Функции мышц и их иннервация
Функции мышц и их иннервация представлены ниже (табл. 2.1).
Таблица 2.1
Функция и иннервация мышц
2.8. Общая симптоматология периферического паралича (пареза)
Периферический паралич, или парез,
представляет собой расстройства произвольных движений, возникающие при поражении периферического двигательного нейрона – самих мотонейронов или их аксонов. Периферический паралич характеризуется следующими признаками:
1) гипотрофия (атрофия) паретичных мышц;
2) снижение мышечного тонуса в паретичных мышцах – мышечная гипотония (атония);
3) утрата или снижение (арефлексия или гипорефлексия) физиологических миотатических и поверхностных рефлексов, которые замыкаются через пораженную часть периферического двигательного нейрона;
4) реакция перерождения в паретичных мышцах;
5) фасцикуляции или фибрилляции в паретичных мышцах.
2.9. Мышечная гипотония при периферическом параличе (парезе)
Под мышечным тонусом
(рис. 2.2) понимают напряжение, в котором находятся мышцы вне активного движения.
Поддержание и перераспределение мышечного тонуса обеспечивает γ-петля. Первым звеном γ-петли являются γ-мотонейроны передних рогов спинного мозга. От γ-мотонейронов идут тонкие γ-волокна к интрафузальным мышечным волокнам, входящим в состав мышечных веретен – мышечных проприоцепторов. Веретена считаются рецепторами натяжения, ответственными за сохранение мышцей постоянной длины. Импульсы, проводимые γ-волокнами, вызывают сокращение интрафузальных мышечных волокон в районах обоих полюсов веретена, тем самым приводя к натяжению его экваториальной части. Это изменение тотчас регистрируется анулоспиральными окончаниями, оплетающими ядерные сумки в веретене (каждое веретено имеет два волокна с ядерной сумкой и четыре – с ядерной цепью). Их потенциал действия увеличивает тонус работающей мышцы.
Возникающий при раздражении анулоспиральных рецепторов нервный импульс идет от веретенных рецепторов по периферическому и центральному отростку клетки спинномозгового ганглия и по заднему корешку поступает в спинной мозг. В результате импульс возвращается преимущественно в тот же сегмент, от которого началась данная γ-петля.
Рис. 2.2.
Схема миотатического рефлекса:
1 – пирамидный путь; 2 – ретикулоспинномозговой путь; 3 – α1-мотонейрон; 4 – α2-мотонейрон; 5 – γ-мотонейрон; 6 – мышечное веретено; 7 – сухожильный рецептор; 8 – чувствительный нейрон
Здесь импульс «переключается» на α-большие и α-малые мотонейроны переднего рога, а также на клетки Реншоу. α-Большие мотонейроны обеспечивают проведение импульсов, вызывающих быстрое (фазическое) сокращение и поддерживающих трофику мышц. α-Малые мотонейроны передают возбуждение на медленно сокращающиеся тонические двигательные единицы. α-Большой мотонейрон отдает коллатераль к клетке Реншоу, а эта клетка, в свою очередь, вновь соединяется с мотонейроном переднего рога, оказывая на него тормозящее действие. Таким образом, основными функциями клеток Реншоу являются функция возвратного торможения α-большого мотонейрона при его слишком сильном возбуждении и функция реципрокной иннервации за счет того, что импульс от клетки Реншоу достигает α-мотонейронов одного сегмента с обеих сторон. В результате по аксонам α-больших и α-малых мотонейронов импульс достигает экстрафузальных мышечных волокон, вызывая их фазическое или тоническое сокращение.
γ-Мотонейроны находятся, в свою очередь, под влиянием волокон, идущих от мотонейронов оральных отделов ствола в составе пирамидного, ретикуло– и вестибулоспинального путей. Таким образом, мышечный тонус может регулироваться как через γ-петлю, так и непосредственно головным мозгом, что важно для каждого произвольного движения. Сокращение интрафузальных мышечных волокон вызывает снижение порога возбудимости рецепторов растяжения. Поэтому влияние γ-мотонейронов через α-малые мотонейроны обеспечивает тонус мышц, а через α-большие мотонейроны – миотатические рефлексы, т. е. фазическое сокращение мышцы в ответ на ее растяжение.
Мышечная гипотония
является следствием «разрыва» соответствующих γ-петель и аксонов от α-малых мотонейронов, идущих в составе поврежденного периферического двигательного нейрона. Мышечная гипотония может быть выявлена при периферическом парезе на основании следующих признаков:
– мышцы дряблые, вялые, тестообразные, их рельеф не контурируется;
– суставы «разболтанные», что проявляется избыточностью движений в них. Так, при мышечной атонии (гипотонии) в руке предплечье свободно прикладывается к плечу, а запястье – к плечевому суставу. На фоне гипотонии в ноге ее можно согнуть в тазобедренном суставе до свободного прикладывания бедра к передней брюшной стенке, а пятки – к ягодице;
– на стороне мышечной гипотонии в ноге отмечается положительный симптом Оршанского, – прижимая колено левой рукой и одновременно отрывая пятку от кушетки правой рукой, врач переразгибает колено пациента до отрывания пятки от кушетки значительно выше, чем в здоровой ноге;
– при выполнении повторных пассивных движений в гипотоничных мышцах отсутствует ощущение сопротивления.
При дифференциальной диагностике мышечной гипотонии следует учитывать, что ее развитие может быть обусловлено не только периферическим парезом, но и другими причинами:
– мышечная гипотония при центральном парезе. Может наблюдаться в случаях его острого развития с последующей трансформацией в мышечную гипертонию;
– мышечная гипотония при заболеваниях экстрапирамидной системы. У этих больных мышечная гипотония сочетается с гиперкинезами – гипотонически-гиперкинетический синдром;
– мышечная гипотония при поражении мозжечка или его проводящих путей. Представляет собой один из характерных признаков мозжечковых расстройств и проявляется целым рядом симптомов;
– мышечная гипотония при заболеваниях с первичным поражением мышц – прогрессирующих мышечных дистрофиях, атрофической миотонии, дермато– и полимиозите и др.;
– физиологическая мышечная гипотония – имеет диффузный характер, сопровождается избыточными движениями в суставах. Определяется у детей и женщин со слабо развитыми мышцами. Избыточные движения в суставах могут создавать ложное впечатление о мышечной гипотонии у спортсменов и акробатов.
2.10. Мышечная гипотрофия при периферическом параличе (парезе)
Мышечная гипотрофия
возникает при периферическом параличе (парезе) вследствие разобщения пораженных мышц с иннервирующими их периферическими мотонейронами. Денервированная мышца перестает получать постоянные трофические импульсы, необходимые для поддержания нормального обмена. В результате в ней наступает перерождение и гибель мышечных волокон с заменой последних жировой и соединительной тканью. Мышечная гипотрофия может быть выявлена при периферическом парезе на основании следующих признаков:
– уменьшение объема мышцы по окружности более чем на 1 см;
– выявление ранних гипотрофий в мышцах языка, кистей, стоп, лопаток, плечевого пояса. На них легко увидеть даже незначительные гипотрофические изменения: сглаженность тенара и гипотенара, «западение» первого межпальцевого промежутка, «выступание» костей на тыле кисти и стопы, изменение формы и конфигурации плечевого сустава, «летающие лопатки» (scapula alatae)
.
Мышечную гипотрофию при периферическом парезе следует дифференцировать с мышечными гипотрофиями при заболеваниях с первичным поражением мышц – прогрессирующих мышечных дистрофиях, атрофической миотонии, поражениях мышц воспалительной природы (дермато– и полимиозит), рефлекторных контрактурах. Кроме того, следует учитывать возможность развития мышечных гипотрофий при эндокринной патологии, кахексии, длительной иммобилизации и в других случаях длительного мышечного бездействия.
2.11. Рефлексы. Строение рефлекторной дуги и рефлекторного кольца
Физиологической основой движения является рефлекс. Это функциональная единица деятельности нервной системы. Впервые понятие «рефлекс» ввел в XVII в. Р. Декарт, который высказал самое общее представление об их существовании.
Рефлекс
(лат.
reflexus
– повернутый назад, отраженный) представляет собой ответную реакцию нервной системы на раздражение, возникающую за счет переноса нервного импульса с афферентных структур на эфферентные.
Рефлексы могут быть охарактеризованы по следующим признакам:
– безусловные (врожденные, передающиеся по наследству) и условные (формирующиеся в процессе индивидуального развития);
– физиологические и патологические (см. ниже);
– замыкающиеся на различных уровнях – спинномозговые, стволовые, подкорковые, корковые;
– поверхностные и глубокие. Поверхностные рефлексы вызываются с кожи и слизистых оболочек, глубокие возникают в ответ на удар неврологическим молоточком по сухожилию или надкостнице (соответственно сухожильные и периостальные).
Рефлекторная дуга
представляет собой анатомо-физиологический субстрат поверхностных рефлексов. Она начинается с рецепторов в коже или слизистых оболочках и включает следующие анатомические структуры: периферический отросток рецепторного нейрона, его тело (в спинномозговых или чувствительных ганглиях черепных нервов), центральный отросток – афферентная часть; вставочный нейрон с его периферическим и центральным отростком – сочетательная часть; мотонейрон в передних рогах спинного мозга или в двигательных ядрах черепных нервов и его аксон, идущий к мышце-исполнителю – эфферентная часть.
Рефлекторное кольцо
имеет, в отличие от рефлекторной дуги, иное строение, поскольку его рецепторы располагаются непосредственно в мышцах и реагируют непосредственно на ее растяжение (
myo
– мышца,
tasis
– растяжение) – миотатические рефлексы. К растяжению мышцы приводит удар неврологическим молоточком по сухожилию или надкостнице, что вызывает растяжение мышечных проприоцепторов – интрафузальных мышечных волокон и мышечных веретен, возникающее в их экваториальной части. Это приводит к раздражению анулоспиральных рецепторов, оплетающих мышечные веретена. От них по тонкому чувствительному периферическому корешку импульс передается на тело и далее на центральный отросток псевдоуниполярного нейрона. В итоге возбуждающий импульс приходит на α-большие и α-малые мотонейроны, а от них по аксону достигает мышц-исполнителей, вызывая соответственно их фазическое (через α-большие) и тоническое (через α-малые) сокращение. Нормальное функционирование рефлекторного кольца всегда связано с γ-петлей, поскольку через нее обеспечивается определенная степень сокращения интрафузальных мышечных волокон, т. е. поддержание мышечного тонуса и предуготованность мышцы к фазическому сокращению.
Проводящие пути среднего мозга и нижележащих структур
Давайте отметим связь проводящих путей среднего мозга с проводящими путями ранее изученных нами структур, чтобы у нас формировалось цельное понятие о проводящих путях ЦНС. В этой подборке я буду «строить» только те проводящие пути, которые напрямую не связаны с черепными нервами, потому что черепные нервы мы будем разбирать отдельным циклом статей.
Красноядерно-спинномозговой тракт
Этот путь начинается от красных ядер мезенцефалона, и, спускаясь вниз, первым делом проходит через мост. Ещё раз обращаем внимание на перекрест Фореля и соединяем средний мозг и мост:
Также мы можем дорисовать всю картину полностью, используя также схемы продолговатого мозга и спинного мозга:
В этой схеме я не нарисовал тело второго нейрона, который располагается в вентральных рогах спинного мозга. Мы нарисуем проводящие пути более подробно в специальном уроке, а пока что запомним все части ЦНС, в которых мы можем видеть руброспинальный тракт.
Не забываем, что руброспинальный путь является одним из основных компонентов эктрапирамидной системы, то есть системы неосознанных движений.
Крыше-спинномозговой тракт
Крыше-спинномозговой, он же — тектоспинальный путь, начинается, соответственно названию, от крыши спинного мозга, там же перекрещивается (перекрест Мейнерта) и далее следует до самого спинного мозга. Отметим связь мезенцефалона и моста по крыше-спинномозговому пути:
По этому рисунку может создаться впечатление, что Варолиев мост содержит ядра тектоспинального пути из-за слишком широкого разреза пучков. Конечно, это просто недочёт моего рисунка, ведь тела вторых нейронов тектоспинального пути располагаются в вентральных рогах спинного мозга.
Включаем в нашу схему ранее изученные части ЦНС и получаем практически завершённый тектоспинальный путь (за исключением тела второго нейрона):
Кортико-спинальный (пирамидный) тракт
В отличие от двух предыдущих путей, мы не видим кортико-спинальный тракт полностью. Значительная часть его проходит через внутреннюю капсулу больших полушарий мозга, а заканчивается он и вовсе в коре больших полушарий. Тем не менее, кортико-спинальный путь проходит красной нитью (ба-дум-тсс) через все изученные нами части ЦНС.
Поэтому мы можем нарисовать примерную схему нижней части этого пути, не забыв про то, что он перекращивается на уровне миеленцефалона, а в спинном мозге он и вовсе разделяется на два пучка — передний и латеральный.
Спинно-таламический тракт
Теперь мы перешли к чувствительным путям. Как вы помните, в спинном мозге был два спинно-таламических тракта — латеральный и передний (также, как и пирамидные тракты). Оба спинно-таламических тракта собирали информацию от чувствительности, воспринимаемой кожей, и передавали её наверх. Волокна спинно-таламических проходят транзитом через миленцефалон, мост и мезенцефалон, направляясь к своей конечной остановке — к таламусу.
Это повзоляет нам примерно нарисовать ход спинно-таламических трактов на нашей схеме:
Медиальная петля
На самом деле, медиальная петля является составной частью проводящего пути глубокой сознательной чувствительности. Слово «глубокий» в данном случае означает «суставно-мышечный», то есть рецепторы этого пути располагаются в мышцах, суставах, фасциях и связках. Глубокой чувствительности противопоставляется поверхностная чувствительность, рецепторы которой располагаются в коже и несут импульс по только что разобранному нами спинно-таламическому тракту.
Полное название этого пути звучит как «ганглио-бульбарно-таламо-кортикальный путь». Предыдущий же, спинно-таламический путь, полностью называется ганглио-спинно-таламо-кортикальный путь. Я специально делаю на этом акцент, потому что многие студенты путаются, называя спино-таламическими трактом медиальную петлю. Перепутать эти пути просто, ведь оба идут в таламус, а после таламуса — в кору головного мозга.
Морфологическое отличие их заключается в том, что тело второго нейрона спинно-таламического тракта находится в спинном мозге, отсюда » ганглио-спинно-таламо-кортикальный путь». Тело второго нейрона пути, участок которого называется медиальной петлёй, находится в продолговатом мозге, также известным как bulbus, отсюда ганглио-бульбо-таламо-кортикальный путь. Проводящие пути с телами нейронов я буду рисовать в других уроках, а пока просто отметим связь мезенцефалона с нижележащими структурами по ганглио-бульбарно-таламо-кортикльному пути, который на большей своей части называется медиальной петлёй:
Латинские термины из этой статьи:
- Tractus rubrospinalis;
- Decussatio tegmenti anterior;
- Tractus tectospinalis;
- Decussatio tegmenti posterior;
- Fasciculus longitudinalis medialis;
- Tractus frontopontinus;
- Tractus occipitotemporopontinus;
- Tractus corticospinalis;
- Tractus corticonuclearis;
- Tractus spinothalamicus;
- Lemniscus medialis;
- Lemniscus lateralis;
- Tractus nigrostriatalis.