Good afternoon, dear readers. Today we will study the midbrain pathways. Before you begin today's topic, be sure to read my tutorials on the spinal cord, medulla oblongata, and pons.
If you follow this order and study the parts of the central nervous system using my articles, you will not have to learn the pathways separately later - you will already know them, because in each new lesson I make connections with the structures from previous lessons.
What is a pathway? A conduction path is a sequence of neurons connected to each other to conduct an impulse in a strictly defined direction. For example, the Gowers pathway, which mediates unconscious proprioceptive sensitivity, consists of two neurons:
- The body of the first neuron is located in the dorsal ganglion, and the axon extends into the muscles/joint capsules/tendons;
- The body of the second neuron is located in the dorsal horn of the spinal cord, its axons extend into the white matter of the spinal cord; then passing through the medulla oblongata and the pons, they enter the cerebellum;
- The body of the third neuron is located in the cortex of the cerebellar vermis.
When we talk about the body and process of a neuron, we understand that in reality we are talking not about one, but about a huge number of neuron bodies (their cluster is called the nucleus) and entire bundles of neuron processes. At the level of each part of the central nervous system there are its own nuclei and its own bundles of fibers. For example, in the last lesson we already studied the nuclei of the midbrain. We will now study the arrangement of the fiber bundles passing through the midbrain. In anatomical textbooks, bundles of fibers that form pathways are called pathways, tracts, or simply pathways.
Midbrain pathways
Like the pathways of other underlying parts of the central nervous system, the pathways of the midbrain are divided into:
- Sensitive (conduct information from receptors, for example, rods/cones, to the central nervous system);
- Motor (conduct information to executive organs located outside the central nervous system, for example, to skeletal muscles);
- Associative (connect areas of the same part of the central nervous system with each other).
However, there is another group of unusual pathways in the midbrain that is associated with the presence of the substantia nigra. The substantia nigra, as you remember from the last guide, synthesizes dopamine and transmits it to the overlying structures of the brain. We will definitely analyze these pathways, but we will start with the more familiar and already known to us motor and sensory pathways of the midbrain.
Motor pathways of the midbrain
Red nuclear spinal tract
When you look at a section of the midbrain, you can't help but notice large, high-contrast structures called red nuclei. The red nuclei are one of the key components of the extrapyramidal motor system - that is, a system of unconscious movements that do not require careful processing in the cerebral cortex.
From the red nuclei down to the spinal cord and, through it, to the skeletal muscles, there is a path called the red nucleus-spinal tract (tractus rubrospinalis). There are also names:
- Monakovsky bundle;
- Rubrospinal tract -
- these are equivalent names for the same conducting path.
We make sure to note the intersection of the rubrospinal fibers, which is located here, at the level of the midbrain. The point of intersection of two rubrospinal tracts has its own name - the anterior tegmental decussation (decussatio tegmenti anterior). According to the author, this anatomical formation is called the cross of Trout.
It looks like a ladder, because in previous lessons I designated the rubrospinal tract that way. I try to keep the same conductive paths in the same color scheme so that it is easier for us to collect them.
Roof-spinal tract
This is another pathway of the extrapyramidal system. The roof-spinal tract (tractus tectospinalis) begins from the sensory nuclei located in the superior and inferior colliculi. As you remember, the subcortical centers of vision and hearing lie there, respectively.
In our case, the term “subcortical” means that there will be fast, immediate processing of visual and auditory signals. After this processing, a switch will be made to the motor neurons of the roof-spinal tract.
Imagine that a metal tray falls out of your field of vision in the room where you are now. You will suddenly hear a sudden, sharp crash. Your body's first reaction will be an involuntary contraction of the skeletal muscles - you will flinch or even jump. It is this reaction that is provided by the roof-spinal tract.
The situation is similar with vision - if some object flashes in your field of vision, you involuntarily turn your head in its direction. If the object's movement is too close to you, you are likely to close your eyes for a split second and cover your face with your hands.
It is these reactions that are subcortical. Let's draw our tectospinal tract, not forgetting to draw the first neurons. Of course, I made a big mistake by not drawing them in the last lesson about the nuclei of the midbrain:
As you can see, the roof-spinal tract also forms a decussation. This section of intersection is called the posterior tegmental decussation (decussatio tegmenti posterior), and according to the author, the decussation of Meynert.
Medial longitudinal fasciculus
The processes of the neuron bodies that form the nuclei of Cajal and Darshkevich are connected into bundles called medial longitudinal bundles (fasciculus longitudinalis medialis). As you remember, this pathway anastomoses with the nuclei of the cranial nerves responsible for eye movement, as well as with the motor fibers of the spinal cord. As a result, the medial longitudinal fasciculus is responsible for combined movements of the head and eyes.
A violation of this function is very similar to what a cheating student looks like, turning his head towards the teacher and his eyes towards his knees and the gadget with cheat sheets.
Frontopontine tract
We will now move to the cerebral peduncles, which are essentially raised, shaped areas of white matter. The fibers of the frontopontine tract (tractus frontopontinus) will be located most medially here. As you remember, in the basilar part of the pons we observed many small nuclei scattered over a large area, which were called the proper nuclei of the pons. The axons of neurons that were grouped into these nuclei formed the middle cerebellar peduncles and were directed, accordingly, to the cerebellum.
It turns out that these axons and nuclei receive information from neurons in the cerebral cortex. The set of fibers that connect the frontal lobe cortex and the pontine nuclei is called the frontopontine tract.
Unlike the tracts we previously studied, the frontopontine tracts do not begin from the nuclei of the midbrain, but pass through it in transit. These pathways are located medial to all other tracts of the cerebral peduncles:
Occipitotemporopontine tract
The occipitotemporopontine tract (tractus occipitotemporopontinus) is completely similar to the frontopontine tract, with the exception that in this case the bodies of the first neurons are not in the frontal gyrus, but in the temporal and occipital gyri. In addition, the bundles of this pathway pass not in the medial, but in the most lateral part of the cerebral peduncles. This is where the differences end:
Corticospinal tract
Finally, we get to the corticospinal tracts (tractus corticospinalis), which in the underlying structures transform into the well-known pyramids.
The corticospinal tracts begin from the cells of the cerebral cortex. These cells lie in the 5th layer of the cortex and are called Betz pyramidal cells. The adjective “pyramidal” is used here because of some similarity between these neurons and pyramids. This is clearly visible when examining microscopic images of Betz pyramidal cells.
The corticospinal tract is a very important evolutionary adaptation. It is the pyramidal system (sometimes the pathways starting from the pyramidal cells are called the pyramidal system) that allows us to make meaningful movements with a preliminary deep, high-quality cortical analysis. For example, when I first tried to tie a surgical knot (it was a Dzhanelidze knot), I had to think for a very long time before each movement. This is where my pyramid system worked at full capacity!
But then, after several thousand repetitions, tying this knot no longer required such a long and thorough analysis on the part of the pyramidal system, and the extrapyramidal system partially came into play.
The fibers of the corticospinal tract, starting from the Betz cells, descend down, passing in transit through the underlying structures of the brain and spinal cord. As you remember, at the level of the medulla oblongata, these tracts cross, after which 80% of the fibers end up on the opposite side. But at the level of the midbrain there are no crossovers, so we simply paint over the part of the cerebral peduncle in which the corticospinal tract lies:
Note 1: the presence of transverse stripes in my drawing is simply the need to somehow visually separate the pathways of the cerebral peduncles from each other, despite the fact that, according to anatomical tradition, all motor pathways should be red, and all sensory pathways should be blue.
Note 2: Topographically, my drawing is not entirely correct because the corticospinal tracts should occupy the largest space of the cerebral peduncle of all the other tracts.
Corticonuclear pathway
The corticonuclear tract (tractus corticonuclearis) is another tract that forms the pyramidal system. However, unlike the corticospinal tract, which innervates the skeletal muscles of the trunk and limbs, the corticonuclear tract innervates the skeletal muscles of the face - chewing and facial muscles, as well as the muscles of the neck.
Sensory pathways of the midbrain
Our midbrain looks too red, doesn't it? A little blue—the traditional color of sensitive pathways—would definitely go a long way here.
Spinothalamic tract
Well known to us from previous lessons, the spinothalamic tract (tractus spinothalamicus) is responsible for transmitting tactile, temperature and pain sensations from skin receptors. At the level of the mesencephalon, the spinothalamic tract does not switch to any nuclei, does not begin and does not end, so we mark the fiber bundle as a horizontal slice:
Medial loop
The medial lemniscus (lemniscus medialis) is the fibers of the pathways of conscious proprioceptive sensitivity. At the level of the spinal cord, these pathways are called the Gaul and Burdach pathways. Further, crossing at the level of the myelencephalon, these paths are connected into one tract, called the medial lemniscus. In the form of a medial loop, these fibers will continue to rise to the thalamus, where they switch to neurons heading to the cerebral cortex.
At the level of the midbrain, no switching occurs; we simply see bundles of fibers:
Lateral loop
The lateral loop (lemniscus lateralis) is the fibers of the pathway that connects the organ of hearing and parts of the central nervous system. At the level of the midbrain, part of the fibers of this pathway are located in the subcortical centers, that is, the lower colliculi of the quadrigeminal plate, and the rest goes directly to the cerebral hemispheres, more precisely, to the cortex of the superior temporal gyrus.
In reality, the lateral loop is not that wide relative to the medial loop, I just had to “flatten” it a little due to lack of space inside the design. In atlases, the lateral loop is depicted as
Routes of dopamine transport. Nigrostriatal pathway
The mesencephalon contains an anatomical formation called the substantia nigra. Neurons of the substantia nigra synthesize dopamine, a hormone and neurotransmitter that transmits impulses in pathways responsible, among other things, for the emergence of a feeling of satisfaction from work and motivation to action. Dopamine is also one of the mediators of the extrapyramidal system.
The disruption of dopamine synthesis in the substantia nigra is undoubtedly associated with a terrible neurological suffering - Parkinson's disease. Another disabling disease, schizophrenia, is also believed to be associated with dopamine metabolism.
In order to deliver dopamine to the overlying structures of the cerebral hemispheres, neurons of the substantia nigra send their processes to the striatum - we will definitely analyze this anatomical formation later. And now we must remember that a significant part of the processes of neurons in the substantia nigra goes upward, to the cerebral hemispheres and carries dopamine there. Three pathways are responsible for this, the most important of which is the nigrostriatal pathway (tractus nigrostriatalis). I schematically marked the course of the nigrostriatal pathway in our illustration:
Text of the book “Clinical diagnostics in neurology”
Chapter 2 DISORDERS OF INVOLUTIONARY AND VOLUNTARY MOVEMENTS
2.1. Types of movements
Movements are divided into involuntary and voluntary. Involuntary movements
are movements that occur regardless of a person’s desire in response to various stimuli. Such reflex movements are inherent in every biological species, are formed at the time of birth, and are inherited. Their anatomical and physiological substrate is a reflex arc or reflex ring, which closes at the level of the spinal cord or brain stem.
Voluntary movements
represent movements of varying degrees of complexity, divided into three groups:
1. Elementary conscious-volitional movements.
They are differentiated motor acts that are voluntary and at the same time have a relatively simple nature - raising an arm or leg, bending them at the joints, etc. These movements are “triggered” on the basis of an efferent impulse formed in the primary cortical field of the motor analyzer (mainly precentral gyrus).
2. Motor praxis.
These are more complex motor acts that are developed in the course of a person’s life on the basis of experience, skill, practice and are reinforced by a stereotype. Programming of such movements (combing one's hair, making the bed, writing, etc.) is carried out in special zones of the cortex (secondary, or associative), and their implementation is also carried out through the primary cortical field of the motor analyzer and its efferent connections.
3. Automated movements
– running, walking, crawling, swimming, etc. – are relatively arbitrary, since their implementation is memorized and carried out as a single motor act. Such movements are the prerogative of the extrapyramidal system and the cerebellum, and the motor fields of the cerebral cortex have a mainly regulatory effect on them.
2.2. Central and peripheral motor neurons of the corticomuscular pathway
To carry out voluntary movement, it is necessary that the motor impulse generated in the corresponding areas of the cerebral cortex be conducted to the skeletal (striated) muscles. This provides the cortico-muscular pathway (tractus corticomuscularis)
, consisting of two parts: the first part is represented by the central motor neuron; the second is a peripheral motor neuron.
Central motor neuron.
According to classical concepts, the bodies of neurons from which the cortico-muscular path originates are located in the anterior central gyrus - the primary cortical field of the motor analyzer.
For a long time it was believed that the central motor neuron originates only from the bodies of the internal large pyramidal neurons (Betz cells), located in the fifth layer of the anterior central gyrus, which predetermined its name - the pyramidal tract. According to recent data, only 27–40% of the axons of the cortical-muscular tract begin from the anterior central gyrus, and only 3–4% directly from Betz cells; about 20% of the fibers of the cortical-muscular tract originate from the somatosensory cortical zone (postcentral gyrus ), and the rest - from the premotor zone, paracentral and other areas of the brain. The axons of these neurons end on the motor neurons of the anterior horns of the spinal cord (corticospinal tract - tractus corticospinalis
) and on the motor nuclei of the cranial nerves (corticonuclear tract -
tractus corticonuclearis
).
Peripheral motor neuron
formed by motor neurons of the anterior horns of the spinal cord and motor nuclei of the cranial nerves, as well as their axons, reaching the performing muscles as part of a number of formations of the nervous system (anterior roots, spinal nerves, plexuses, peripheral or cranial nerves).
2.3. Anatomy of the corticospinal and corticonuclear tracts
Corticospinal tract
starts mainly from the upper 2/3 of the anterior central gyrus and paracentral lobule; some of the axons originate in the adjacent premotor area, in the posterior central gyrus and superior parietal lobule (Fig. 2.1).
Rice. 2.1.
Pyramid path diagram:
1 – lateral (crossed) pyramidal bundle; 2 – straight (uncrossed) pyramidal bundle; 3 – cortical-nuclear fibers; 4 – projection of the motor sphere in the anterior central gyrus of the cerebral cortex
In the cortex of the anterior central gyrus, pyramidal neurons are localized according to the “motor homunculus” rule (Penfield scheme). This means that in the uppermost parts of the anterior central gyrus there are neurons that begin the efferent pathways for the leg muscles: in the paracentral lobule there are neurons that provide movement of the foot muscles, and in the upper parts of the anterior central gyrus there is a sequential somatotopic projection for the muscles of the lower leg and thigh. Next, neurons are located in succession, giving rise to efferent nerve pathways to the muscles of the trunk. The middle third of the anterior central gyrus is occupied by neurons that provide innervation to the arm muscles. The area of somatotopic projection zones in the anterior central cortex is proportional to the complexity of movements performed by a particular muscle group, with the largest area occupied by the somatotopic projection of the muscles of the hand (especially the thenar muscles).
Axons from the above areas of the anterior central gyrus pass through the corona radiata, the anterior 2/3 of the posterior femur of the internal capsule and then follow along the ventral surface of the brain stem. At the border between the medulla oblongata and the spinal cord, 80% of the fibers of the corticospinal tract pass to the opposite side, forming a pyramidal decussation (decussatio pyramidum)
.
The crossed fibers are sent to the lateral cord of the spinal cord, occupying its posteromedial section in the form of the lateral corticospinal tract (tractus corticospinalis lateralis)
.
This path provides voluntary movements of both the muscles of the trunk and limbs. About 20% of the fibers remain uncrossed, forming the anterior corticospinal tract (tractus corticospinalis anterior)
. This pathway provides voluntary movements mainly in the muscles of the trunk and neck. The fibers of the lateral corticospinal tract are located in the spinal cord in accordance with the Auerbach–Flatau law and at the same time switch segment by segment to the motor neurons of the anterior horns of the spinal cord.
Corticonuclear pathway
begins predominantly from the lower third of the anterior central gyrus and the adjacent premotor area. A clear somatotopic projection of the muscles innervated by the cranial nerves is also determined here, and in the lowest part of the precentral gyrus there are neurons for the muscles of the pharynx, larynx, soft palate, tongue, masticatory and facial muscles. The axons of the corticonuclear tract pass through the corona radiata, the genu of the internal capsule, and the brainstem. Here its fibers make a partial supranuclear decussation, ending on the motor nuclei of the cranial nerves of their own and the opposite side. The exception is the part of the corticonuclear tract that ends on the nucleus of the XII and the lower part of the nucleus of the VII nerve - these fibers make a complete supranuclear decussation and therefore end on the corresponding motor nuclei (XII, the lower half of VII) only on the opposite side.
2.4. Plegia and paresis
Plegia, or paralysis,
– complete loss of voluntary movements in one or another muscle group.
Paresis
– partial loss of voluntary movements in one or another muscle group, characterized by a decrease in the strength and range of active movements in the affected muscles.
Depending on the prevalence, the following types of paralysis and paresis are distinguished:
– monoplegia, or monoparesis, – plegia or paresis is determined only in the muscles of one arm or leg;
– hemiplegia, or hemiparesis, – plegia or paresis affects the muscles of both the arms and legs on one or opposite (alternating hemiplegia or hemiparesis) sides of the body;
– paraplegia, or paraparesis, – plegia or paresis is determined only in the muscles of both arms or legs (upper or lower paraplegia or paraparesis, respectively);
– triplegia, or triparesis, – plegia or paresis involves three limbs;
– tetraplegia, or tetraparesis, – plegia or paresis affects both arms and legs.
2.5. Common clinical signs of movement disorders
Plegia (paresis) can be detected during a clinical examination based on the following signs:
1) decrease in the range of active movements and (or) muscle strength;
2) changes in muscle tone;
3) atrophy or hypotrophy of paretic muscles;
4) fasciculations and fibrillation of the affected muscles;
5) increase or decrease (up to absence) of physiological reflexes that are closed on paretic muscles;
6) the presence of pathological reflexes;
7) the presence of protective reflexes and pathological synkinesis. Depending on the type of plegia or paresis - peripheral or central - the various above symptoms form the clinical core of peripheral or central paralysis.
2.6. Methods for studying muscle strength of various muscle groups
Muscle strength is studied in parallel with active movements, since their volume does not decrease with mild paresis. The strength of the hand muscles is determined with a dynamometer. When studying the strength of other muscles, the manual method is used in two modifications.
During the first modification, the doctor interferes with the performance of an active movement, determines and compares the resistance force in the corresponding muscles on the left and right. So, for example, the doctor asks the patient to bend his arm at the elbow joint on the left and actively prevents this bending. Then the strength of the biceps brachii muscle of the right hand is also determined, and the strength of the active movement on the left and right is compared.
Another modification is more often used: the patient is asked to perform an active movement without providing resistance. Next, the patient holds the arm or leg in this position with maximum force, and the doctor tries to make a movement in the opposite direction. At the same time, he evaluates and compares on the left and right the degree of effort that is required for this. For example, the strength of the biceps brachii muscle is determined by trying to straighten an arm that is already bent at the elbow joint, first on the left, then on the right.
2.7. Muscle functions and their innervation
The functions of the muscles and their innervation are presented below (Table 2.1).
Table 2.1
Muscle function and innervation
2.8. General symptomatology of peripheral paralysis (paresis)
Peripheral paralysis, or paresis,
is a disorder of voluntary movements that occurs when a peripheral motor neuron is damaged - the motor neurons themselves or their axons. Peripheral paralysis is characterized by the following symptoms:
1) hypotrophy (atrophy) of paretic muscles;
2) decreased muscle tone in paretic muscles - muscle hypotonia (atony);
3) loss or decrease (areflexia or hyporeflexia) of physiological myotatic and superficial reflexes, which are closed through the affected part of the peripheral motor neuron;
4) reaction of degeneration in paretic muscles;
5) fasciculations or fibrillations in paretic muscles.
2.9. Muscular hypotonia in peripheral paralysis (paresis)
Under muscle tone
(Fig. 2.2) understand the tension in which the muscles are located outside of active movement.
The maintenance and redistribution of muscle tone is ensured by the γ-loop. The first link of the γ-loop is the γ-motoneurons of the anterior horns of the spinal cord. Thin γ-fibers go from γ-motoneurons to intrafusal muscle fibers, which are part of the muscle spindles - muscle proprioceptors. The spindles are considered to be tension receptors responsible for maintaining the muscle at a constant length. Impulses conducted by γ-fibers cause contraction of intrafusal muscle fibers in the areas of both poles of the spindle, thereby leading to tension in its equatorial part. This change is immediately registered by anulospiral endings entwining the nuclear bags in the spindle (each spindle has two fibers with a nuclear bag and four with a nuclear chain). Their action potential increases the tone of the working muscle.
The nerve impulse arising from irritation of the anulospiral receptors travels from the spindle receptors along the peripheral and central processes of the dorsal ganglion cell and enters the spinal cord along the dorsal root. As a result, the impulse returns predominantly to the same segment from which this γ-loop began.
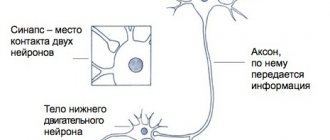
Rice. 2.2.
Diagram of the myotatic reflex:
1 – pyramidal path; 2 – reticulospinal tract; 3 – α1-motoneuron; 4 – α2-motoneuron; 5 – γ-motoneuron; 6 – muscle spindle; 7 – tendon receptor; 8 – sensitive neuron
Here the impulse “switches” to the α-major and α-small motor neurons of the anterior horn, as well as to Renshaw cells. α-Large motor neurons provide impulses that cause rapid (phasic) contraction and maintain muscle trophism. α-Small motor neurons transmit excitation to slow-twitch tonic motor units. The α-Large motor neuron sends a collateral to the Renshaw cell, and this cell, in turn, reconnects with the anterior horn motor neuron, exerting an inhibitory effect on it. Thus, the main functions of Renshaw cells are the function of reciprocal inhibition of the α-large motor neuron when it is too excited and the function of reciprocal innervation due to the fact that the impulse from the Renshaw cell reaches the α-motoneurons of one segment on both sides. As a result, along the axons of the α-large and α-small motor neurons, the impulse reaches the extrafusal muscle fibers, causing their phasic or tonic contraction.
γ-Motoneurons are, in turn, under the influence of fibers coming from the motor neurons of the oral sections of the trunk as part of the pyramidal, reticulo- and vestibulospinal tracts. Thus, muscle tone can be regulated both through the γ-loop and directly by the brain, which is important for every voluntary movement. Contraction of intrafusal muscle fibers causes a decrease in the threshold of excitability of stretch receptors. Therefore, the influence of γ-motoneurons through α-small motor neurons provides muscle tone, and through α-large motor neurons - myotatic reflexes, i.e., phasic contraction of the muscle in response to its stretching.
Muscular hypotonia
is a consequence of the “break” of the corresponding γ-loops and axons from α-small motor neurons running as part of the damaged peripheral motor neuron. Muscular hypotonia can be detected in peripheral paresis based on the following signs:
– muscles are flabby, sluggish, pasty, their relief is not contoured;
– joints are “loose”, which is manifested by excessive movements in them. So, with muscle atony (hypotonia) in the hand, the forearm is freely applied to the shoulder, and the wrist is freely applied to the shoulder joint. Against the background of hypotension in the leg, it can be bent at the hip joint until the thigh is freely attached to the anterior abdominal wall, and the heels are attached to the buttock;
- on the side of muscle hypotonia in the leg, a positive Orshansky sign is noted - pressing the knee with the left hand and at the same time lifting the heel off the couch with the right hand, the doctor hyperextends the patient’s knee until the heel lifts off the couch significantly higher than in the healthy leg;
– when performing repeated passive movements in hypotonic muscles there is no feeling of resistance.
When differentially diagnosing muscle hypotonia, it should be taken into account that its development may be due not only to peripheral paresis, but also to other reasons:
– muscle hypotonia with central paresis. It can be observed in cases of its acute development with subsequent transformation into muscle hypertension;
– muscle hypotonia in diseases of the extrapyramidal system. In these patients, muscle hypotonia is combined with hyperkinesis - hypotonic-hyperkinetic syndrome;
– muscle hypotonia with damage to the cerebellum or its pathways. It is one of the characteristic signs of cerebellar disorders and is manifested by a number of symptoms;
– muscle hypotonia in diseases with primary muscle damage – progressive muscular dystrophies, atrophic myotonia, dermato- and polymyositis, etc.;
– physiological muscle hypotonia – has a diffuse nature, accompanied by excessive movements in the joints. It is determined in children and women with poorly developed muscles. Excessive joint movements can create a false impression of muscle hypotonia in athletes and acrobats.
2.10. Muscle wasting in peripheral paralysis (paresis)
Muscle wasting
occurs with peripheral paralysis (paresis) due to the disconnection of the affected muscles from the peripheral motor neurons that innervate them. A denervated muscle ceases to receive constant trophic impulses necessary to maintain normal metabolism. As a result, degeneration and death of muscle fibers occurs in it, with the latter being replaced by fatty and connective tissue. Muscle wasting can be detected in peripheral paresis based on the following signs:
– reduction in muscle volume around the circumference by more than 1 cm;
– detection of early malnutrition in the muscles of the tongue, hands, feet, shoulder blades, and shoulder girdle. It is easy to see even minor hypotrophic changes on them: smoothness of the thenar and hypothenar, “recession” of the first interdigital space, “protrusion” of the bones on the back of the hand and foot, changes in the shape and configuration of the shoulder joint, “flying shoulder blades” (scapula alatae)
.
Muscle wasting in peripheral paresis should be differentiated from muscle wasting in diseases with primary muscle damage - progressive muscular dystrophy, atrophic myotonia, muscle damage of an inflammatory nature (dermato- and polymyositis), reflex contractures. In addition, the possibility of developing muscle wasting due to endocrine pathology, cachexia, prolonged immobilization and other cases of prolonged muscle inactivity should be taken into account.
2.11. Reflexes. The structure of the reflex arc and reflex ring
The physiological basis of movement is a reflex. It is the functional unit of activity of the nervous system. The concept of “reflex” was first introduced in the 17th century. R. Descartes, who expressed the most general idea about their existence.
Reflex
(Latin
reflexus
- turned back, reflected) is a response of the nervous system to irritation that occurs due to the transfer of a nerve impulse from afferent structures to efferent ones.
Reflexes can be characterized by the following characteristics:
– unconditional (congenital, inherited) and conditional (formed in the process of individual development);
– physiological and pathological (see below);
– closing at various levels – spinal, brainstem, subcortical, cortical;
– superficial and deep. Superficial reflexes are caused from the skin and mucous membranes, deep reflexes occur in response to a blow with a neurological hammer on the tendon or periosteum (tendon and periosteal, respectively).
Reflex arc
is an anatomical and physiological substrate of surface reflexes. It begins with receptors in the skin or mucous membranes and includes the following anatomical structures: the peripheral process of the receptor neuron, its body (in the spinal or sensory ganglia of the cranial nerves), the central process - the afferent part; interneuron with its peripheral and central processes – the combining part; a motor neuron in the anterior horns of the spinal cord or in the motor nuclei of the cranial nerves and its axon going to the executive muscle - the efferent part.
Reflex ring
has, in contrast to the reflex arc, a different structure, since its receptors are located directly in the muscles and react directly to its stretching (
myo
- muscle,
tasis
- stretching) - myotatic reflexes. A muscle stretch is caused by a blow to the tendon or periosteum with a neurological hammer, which causes stretching of the muscle proprioceptors - intrafusal muscle fibers and muscle spindles, occurring in their equatorial part. This leads to irritation of the anulospiral receptors intertwining the muscle spindles. From them, along a thin sensitive peripheral root, the impulse is transmitted to the body and further to the central process of the pseudounipolar neuron. As a result, the exciting impulse arrives at the α-major and α-small motor neurons, and from them along the axon reaches the performing muscles, causing their phasic (through the α-large) and tonic (through the α-small) contraction, respectively. The normal functioning of the reflex ring is always associated with the γ-loop, since through it a certain degree of contraction of intrafusal muscle fibers is ensured, i.e., maintaining muscle tone and preparing the muscle for phasic contraction.
Conducting pathways of the midbrain and underlying structures
Let's note the connection of the midbrain pathways with the pathways of the structures we previously studied, so that we can form a complete concept of the pathways of the central nervous system. In this collection, I will “build” only those pathways that are not directly connected to the cranial nerves, because we will examine the cranial nerves in a separate series of articles.
Red nuclear spinal tract
This path starts from the red nuclei of the mesencephalon, and, going down, first passes through the bridge. Once again we pay attention to the Trout cross and connect the midbrain and the pons:
We can also complete the whole picture using diagrams of the medulla oblongata and spinal cord:
In this diagram, I did not draw the body of the second neuron, which is located in the ventral horns of the spinal cord. We will draw the pathways in more detail in a special lesson, but for now we will remember all the parts of the central nervous system in which we can see the rubrospinal tract.
Do not forget that the rubrospinal tract is one of the main components of the extrapyramidal system, that is, the system of unconscious movements.
Roof-spinal tract
The roof-spinal cord, also known as the tectospinal tract, begins, according to its name, from the roof of the spinal cord, crosses there (Meynert's cross) and then follows all the way to the spinal cord. Let us note the connection between the mesencephalon and the bridge along the roof-spinal tract:
This figure may give the impression that the pons contains nuclei of the tectospinal tract due to the too wide section of the fascicles. Of course, this is just a flaw in my drawing, because the bodies of the second neurons of the tectospinal tract are located in the ventral horns of the spinal cord.
We include the previously studied parts of the central nervous system into our diagram and obtain an almost complete tectospinal tract (with the exception of the body of the second neuron):
Corticospinal (pyramidal) tract
Unlike the previous two tracts, we do not see the entire corticospinal tract. A significant part of it passes through the internal capsule of the cerebral hemispheres, and it ends in the cerebral cortex. However, the corticospinal tract runs like a red thread (ba-dum-shh) through all the parts of the central nervous system we have studied.
Therefore, we can draw an approximate diagram of the lower part of this path, not forgetting that it is repainted at the level of the myelencephalon, and in the spinal cord it is completely divided into two bundles - anterior and lateral.
Spinothalamic tract
We have now moved on to the sensitive pathways. As you remember, the spinal cord had two spinothalamic tracts - lateral and anterior (as well as the pyramidal tracts). Both spinothalamic tracts collected information from the sensations perceived by the skin and transmitted it upward. The spinothalamic fibers transit through the mylencephalon, pons and mesencephalon, heading to their final stop - the thalamus.
This allows us to roughly draw the course of the spinothalamic tracts on our diagram:
Medial loop
In fact, the medial lemniscus is part of the deep conscious sensitivity pathway. The word “deep” in this case means “articular-muscular”, that is, the receptors of this pathway are located in the muscles, joints, fascia and ligaments. Deep sensitivity is contrasted with superficial sensitivity, the receptors of which are located in the skin and carry an impulse along the spinothalamic tract we just discussed.
The full name of this pathway is “ganglio-bulbar-thalamo-cortical pathway.” The previous one, the spinothalamic tract, is completely called the ganglio-dorsal-thalamo-cortical tract. I specifically emphasize this because many students get confused when they call the medial lemniscus the spinothalamic tract. It is easy to confuse these pathways, because both go to the thalamus, and after the thalamus - to the cerebral cortex.
Their morphological difference lies in the fact that the body of the second neuron of the spinothalamic tract is located in the spinal cord, hence the “ganglio-spinal-thalamo-cortical tract.” The body of the second neuron in the tract, a portion of which is called the medial lemniscus, is located in the medulla oblongata, also known as the bulbus, hence the gangliobulbothalamocortical tract. I will draw the pathways with neuron bodies in other lessons, but for now let’s just note the connection of the mesencephalon with the underlying structures along the ganglio-bulbar-thalamo-cortical pathway, which for the most part is called the medial lemniscus:
Latin terms from this article:
- Tractus rubrospinalis;
- Decussatio tegmenti anterior;
- Tractus tectospinalis;
- Decussatio tegmenti posterior;
- Fasciculus longitudinalis medialis;
- Tractus frontopontinus;
- Tractus occipitotemporopontinus;
- Tractus corticospinalis;
- Tractus corticonuclearis;
- Tractus spinothalamicus;
- Lemniscus medialis;
- Lemniscus lateralis;
- Tractus nigrostriatalis.