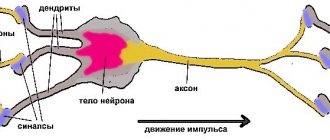
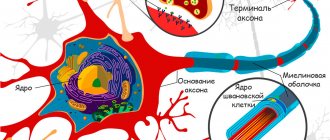
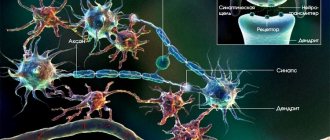
When neuroscientists talk about dendrites—the parts of neurons typically used to receive information from other neurons—they think of Purinier cells.
They form an extensive specialized network that interacts with the ventricular minutes to initiate ventricular contraction.
Computational studies suggest that cellular sources of arrhythmia, such as early afterdepolarization, may occur due to their unique electrophysiological features.
Activity of Purkinje cells in the cerebellar cortex
Purkinje cells provide output from the cerebellar cortex. Their dendrites contain many excitatory synapses from parallel fibers of granule cells (Fig. 40.10 and Fig. 40.11). In addition, each Purkinje cell receives extensive excitatory connections from a liana-shaped fiber, the processes of which intertwine their dendritic tree, forming numerous synaptic contacts. Signals from mossy fibers produce single action potentials (simple spikes) in Purkinje cells, while the responses of these cells to signals from an individual liana fiber are rhythmic discharges (complex spikes) (Fig. 40.12). Because the frequency of complex adhesions is low, they do not increase the mean firing rate of Purkinje cells, but do appear to alter their sensitivity to mossy fiber inputs. Such changes may be long-lasting and therefore must play a role in motor learning.
Purkinje cells receive inhibitory signals from basket cells and stellate cells (Fig. 40.10 and Fig. 40.11); as a result, inhibitory postsynaptic potentials are generated in them (Fig. 40.12). Purkinje cell axons descend through the granular layer into the white matter of the cerebellum (Fig. 40.11). Most of these axons terminate in one of the deep cerebellar nuclei. Some Purkinje cells in the vestibulocerebellum project to the lateral vestibular nucleus. Their discharge leads to inhibition of the activity of neurons in the deep cerebellar nuclei and lateral vestibular nucleus. GABA is released from Purkinje cells as an inhibitory neurotransmitter.
In Fig. Figure 40.13 shows some cerebellar neural networks formed by afferent fibers, interneurons and Purkinje cells. Mossy fibers cause activation of granule cells and Purkinje cells, as well as, through basket and stellate cells, direct (feedforward) inhibition of the latter (Fig. 40.13a). Liana-shaped fibers create powerful excitation of Purkinje cells (Fig. 40.13, c). Inputs from mossy fibers are regulated by inhibitory feedback to Golgi cells (Fig. 40.13c). Complex interactions occur between mossy and liana-like fibers (Fig. 40.13e). At first glance, the inhibitory nature of the output from the cerebellar cortex may seem paradoxical. However, it must be taken into account that all the pathways included in it give collaterals to the deep nuclei of the cerebellum. Therefore, the high activity of neurons in these nuclei is not surprising. The cerebellar cortex must modulate this activity. Thus, the real output from the cerebellum is through projections from the deep nuclei.
Story
For the first time, as you might guess, Purkinje cells were seen by a man named Purkinje. Or Purkina, as purists like to say. The Czech Jan Evangelista Purkinje was an amazing person, an encyclopedist, corresponded with Goethe, was a member of the Order of the Illuminati, discovered twilight vision, became one of the founders of fingerprinting and created a prototype of a movie camera, despite the fact that he was an older contemporary of Pushkin. He lived a long life and studied anatomy for just over two decades. In 1837, he described “ganglionic bodies” - Purkinje cells.
Jan Evangelista Purkinje
The most famous image of these branched cells was obtained by another great scientist - the 1906 Nobel laureate, Santiago Ramon y Cajal. His famous drawing, reproduced in all textbooks, depicts Purkinje cells and deeper granular cells in the cerebellum of a pigeon.
Purkinje cells (A) and granule cells (B) of the pigeon cerebellum. Drawing of Santiago Ramon y Cajal
Interneuronal connections of the cerebellum
Cerebellar neurons do not have direct output to spinal motor neurons, but act on them through cortical-stem motor centers.
Piriform neurons (Purkinje cells) form the middle layer of the cerebellar cortex and are the main functional unit with a pronounced integrative function. The structural basis of this function is the numerous branching dendrites of piriform cells.
Purkinje cells are the only efferent neurons of the cerebellar cortex and directly connect it with the intracerebellar and vestibular nuclei.
The main cerebellar afferents are liana and mossy fibers.
Liana-like fibers. The main source of liana-shaped fibers are neurons of the inferior olive of the medulla oblongata. Information comes to them from muscle and skin receptors, as well as from the motor cortex of the cerebral hemispheres. Each liana-shaped fiber establishes synaptic contacts with dendrites (transmitter aspartate) of one Purkinje cell. The number of synapses can reach three hundred, which explains the exceptionally strong excitatory effect of this afferent input (a large EPSP, at the top of which 3-5 action potentials are formed).
In addition, these fibers have a weak inhibitory effect on Purkinje cells through the basket and stellate cells of the superficial layer of the cerebellar cortex.
Mossy fibers send information to the cerebellar cortex from the cerebral cortex, proprioceptors of the musculoskeletal system, vestibular receptors and the reticular formation.
Mossy fibers form excitatory synapses on the dendrites of granule cells in the inner layer of the cerebellar cortex. Through the axons of granule cells, this afferent input has a direct excitatory effect (the mediator glutamate) and an indirect inhibitory effect on Purkinje cells. The activity of granule cells is regulated through the inhibitory Golgi cells of the inner layer of the cerebellar cortex according to the type of recurrent inhibition (GABA mediator).
In the cerebellum, the inhibitory nature of control dominates, which is explained by the fact that Purkinje cells, being inhibitory neurons, convert excitatory signals at the input into inhibitory signals at the output.
Thus, the efferent influence of the cerebellar cortex on the subsequent neural link is carried out through the mechanism of restraining neuronal activity.
When studying the connections of the cerebellar cortex with its nuclei, three symmetrical vertical zones were identified.
● Purkinje cells of the medial vermiform zone, as well as the flocculus and nodule, project to the tent nuclei.
The neurons of these nuclei are connected to the motor centers of the trunk, from which the vestibulo-, rubro- and reticulospinal tracts that regulate muscle tone go to the spinal motor centers.
● Purkinje cells of the intermediate zone of the cerebellum project to the globular and cortical nuclei.
The axons of the neurons of these nuclei go to the red nucleus of the midbrain, from which the rubrospinal tract begins, stimulating the tone of the flexor muscles through the spinal motor centers.
● Purkinje cells of the lateral zone of the cerebellar cortex project to the dentate nuclei of the cerebellum, from where pathways are directed to the ventrolateral nucleus of the thalamus, the neurons of which project to the motor areas of the cerebral cortex.
So, three paired cerebellar nuclei form the main efferent outputs of the cerebellum to the stem and cortical motor centers.
The neurons of these nuclei have high background activity. They are under the inhibitory influence of Purkinje cells and the excitatory influence of afferent inputs to the cerebellum, which enter these nuclei through collateral branches.
Fig.1. Scheme of the main interneuron connections of the cerebellum:
CP – Purkinje cells; kG – Golgi cells; kz – granule cells; kK – basket cells; zk – stellate cells. “+” - exciting and “-” inhibitory influences.
What are Purkinje neurons?
Purkinje cells or neurons are named after the Czech-born anatomist, physiologist and botanist Jan Evangelista Purkinje, who discovered these elements. These large cells are found in all invertebrate animals. , are a type of GABAergic neuron and constitute the functional units of the cerebellum.
After its discovery, there were many researchers who tried to decipher the mysteries of this neuron. Renowned scientists Camillo Golgi and Santiago Ramon y Cajal spent years of their lives studying these chambers. Thanks to these studies, we now have almost absolute knowledge about the anatomy and structure of Purkinje neurons, as well as the details and specific functions of these.
Although they are mainly found in the cerebellar cortex, forming the Purkinje layer between the molecular layer and the granular layer, they can also be found in the myocardium, that is, the muscular part of the heart.
Functions of the cerebellum
The functions of the cerebellum form its three main influences on the body: on the motor apparatus, afferent systems and the autonomic system.
► The motor functions of the cerebellum include the regulation of muscle tone, posture and balance, coordination of posture and movement performed, as well as programming goal-directed movements.
●Regulation of muscle tone, posture and balance
carried out mainly by the ancient cerebellum and partly by the old cerebellum.
Receiving and processing impulses from vestibular receptors, proprioceptors of the movement apparatus, skin, visual and auditory receptors, the cerebellum is able to assess the state of the muscles, the position of the body in space and through the nuclei of the tent, using the vestibulo-, reticulo- and rubrospinal tracts, redistribute muscle tone, change posture body and maintain balance.
Impaired balance is the most characteristic symptom of damage to the ancient cerebellum.
●Coordination of posture and targeted movement performed
carried out by the old and new cerebellum. The cortex of this part of the cerebellum receives impulses from the receptors of the movement apparatus, as well as impulses from the motor cortex. By analyzing information about the movement program (from the motor cortex) and the execution of the movement (from proprioceptors), the cerebellum is able, through its intermediate nuclei, which have access to the red nucleus and the motor cortex, to coordinate the posture and the targeted movement being performed, as well as correct the direction of movement.
Impaired motor coordination (dynamic ataxia) is the most characteristic symptom of dysfunction of the intermediate zone of the cerebellum.
●The cerebellum is involved in programming goal-directed movements
. This function is carried out by that part of the cerebellar hemispheres that is included in the lateral zone.
Functions
The main function of Purkinje cells is the regulation of motor activity. There are about 26 million such cells in the cerebellar cortex. Their formation is completed at the age of 8 years. Young children, whose cell development process has not yet completed, move awkwardly and look clumsy. Training stimulates their motor activity and increases the speed of physical development. In children who engage in dancing, ballet or sports that require the reproduction of precise and clear small movements (gymnastics, martial arts), the cerebellum is most developed.
When a person learns to perform target movements (ride a bicycle, hit a basketball hoop, control a computer mouse), he is not immediately able to accurately perform the intended actions. This happens because the muscles tense in the wrong place and at the wrong time. During training, the brain improves body control, and after several not very successful attempts, a person masters a motor skill.
Purkinje cells independently generate a predictive impulse, which reflects information about the upcoming movement. In response, they receive information about errors and inaccuracies that occurred during the execution of the movement. By comparing information, the cerebellum corrects it. When developing a program of motor activity, a complex movement is divided into separate fragments. A specific group of Purkinje cells is responsible for the execution of each fragment.
In modern neurobiology, the area of Purkinje cells is designated as a zone where motor memory is formed and stored - information about all completed movements. Memory contains data about motor actions performed by a person every day, and about those that are improved in the process of further motor learning.
Purkinje cells
The cortex of this part of the cerebellum receives impulses from the association zones of the cerebral cortex through the pontine nuclei. In the neocerebellar cortex, it is processed into a movement program, which, through the dentate nucleus of the cerebellum and the ventrolateral nucleus of the thalamus, enters the premotor and motor cortex of the cerebral hemispheres.
There, this program receives further processing and is implemented through the pyramidal and extrapyramidal systems as a complex, purposeful movement.
Fig.2. Scheme of cerebellar functions: a - regulation of tone, posture and balance; b – coordination of posture and targeted movements; c – programming of voluntary movements.
Control and correction of slower programmed movements is carried out by the cerebellum based on feedback afferentation from proprioceptors, as well as from vestibular, visual and tactile receptors.
Correction of fast movements occurs in a short time by changing the program in the lateral cerebellum, a network based on learning and previous experience.
► Afferent function of the cerebellum. The cerebellum has complex bidirectional connections with sensory systems. The mechanisms of the cerebellum’s influence on sensory functions are associated with its influence on the efferent control of the activity of the receptor apparatus and the switching center in sensory systems.
In the implementation of the influence of the cerebellum on the afferent systems of the body, projections of the cerebellar nuclei onto the specific and nonspecific nuclei of the thalamus as the main switching center in sensory systems play an important role.
► The role of the cerebellum in the regulation of autonomic functions. The cerebellum is the highest autonomic center. Autonomic functions are influenced by the old and ancient cerebellum (mainly the vermis), which receives part of the impulse from interoreceptors.
The cerebellum exerts efferent influences on the autonomic sphere through the nuclei of the tent. They are realized through the nuclei of the reticular formation of the trunk and can be excitatory, inhibitory and mixed. The specific mechanisms for processing interoceptive impulses by the cerebellum are unknown.
Cells of the third type
Histologists have identified several types of cells in the terminal sections of the conduction system of the heart. According to the classification considered here, cells of the third type will have a similar structure to those that make up the Purkinje fibers in the heart. They are more voluminous compared to other pacemakers, long and wide. The thickness of the myofibrils is not the same in all parts of the fiber, but the sum of all contractile elements is greater than in a normal cardiomyocyte.
Important Pallister-killiam syndrome symptoms, causes, treatment
Now we can compare cells of the third type with those that make up the Purkinje fibers. The histology (preparation obtained from tissue at the apex of the heart) of these elements differs significantly. The nucleus has an almost rectangular shape, and the contractile fibers are rather poorly developed, have many branches and are interconnected. In addition, they are not clearly oriented along the length of the cell and are located at large intervals. A meager number of organelles that are located around the myofibrils.
Differences in the frequency of generated impulses and the speed of their conduction require a phylogenetically developed mechanism for synchronizing the contraction process in all parts of the heart.
Cerebellum and motor coordination disorder
Human motor skills are characterized by the accuracy of purposeful movements, which is ensured by the proportional work of many muscle groups, controlled voluntarily and automatically. This complex multifunctional system is implemented by a multineural coordinating apparatus that controls the balance of the body, stabilizes the center of gravity, regulates the tone and coordinated various activities of the muscles.
The center of coordination of movements is the cerebellum; functionally, it contains the body of the cerebellum, consisting of two hemispheres, the vermis and three pairs of peduncles.
In the implementation of voluntary movement, the main role of the cerebellum is to coordinate the fast and slow components of the motor act.
This becomes possible thanks to the bilateral connections of the cerebellum with the muscles and cerebral cortex. The cerebellum receives afferent impulses from all receptors irritated during movement (from proprioceptors, vestibular, visual, auditory, etc.).
Receiving information about the state of the motor system, the cerebellum influences the red nucleus of the brain, which sends impulses to the γ-motoneurons of the spinal cord, which regulate muscle tone. In addition, part of the afferent impulses through the cerebellum enters the cerebral cortex of the motor zone (precentral and frontal gyri).
The main function of the cerebellum is carried out at a subconscious level.
Efferent impulses from the cerebellar nuclei regulate proprioceptive stretch reflexes. During muscle contraction, the proprioceptor of the synergist muscles and antagonist muscles are excited. Normally, the transformation of voluntary movement into a complex reflex does not occur due to the inhibitory influence of cerebellar impulses. Therefore, when the cerebellum is damaged, disinhibition of segmental proprioceptive reflexes is manifested by movements of the limbs like ataxia.
The cerebellum has many afferent and efferent connections.
Posterior spinocerebellar tract (Flexig's tract). The first neuron is located in the spinal ganglion; the axon in the dorsal root, through the dorsal horn, approaches the cells of Clark's column. The fibers of these second neurons are directed to the outer layers of the posterior part of the lateral funiculus on their side, ascend along the entire spinal cord and, at the level of the medulla oblongata, as part of the inferior cerebellar peduncle, enter the cerebellar vermis.
This pathway is referred to as the posterior spinocerebellar tract. In the cortex of the cerebellar vermis there is a third neuron that contacts the piriform neurons of the cerebellar hemisphere cortex. The axons of the latter go to the dentate nucleus. The fibers of this fifth neuron are part of the superior cerebellar peduncle.
The right and left superior cerebellar peduncles cross - Wernicke's cross - and end at the red nucleus cells of the opposite side. The axons of the cells of the red nucleus are immediately directed to the opposite side of the midbrain and form a ventral decussation in the tegmentum of the midbrain - the decussation of Forel, pass as part of the lateral cord of the spinal cord, and reach the cells of the anterior horns.
The set of axons of the cells of the red nucleus is called the Monakov bundle.
Govers' anterior spinocerebellar tract. The first neuron is located in the spinal ganglion, the second neuron is a cell of the dorsal horn, but its axons move to the opposite side and go up the spinal cord, in the anterior part of the lateral cord, pass through the medulla oblongata, the pons of the brain, at the level of the superior medullary velum they pass to the opposite side side and as part of the superior cerebellar peduncle reach the cells of the cerebellar nuclei.
The cerebellum receives afferent proprioceptive impulses not only along the Flexig and Gowers pathways, they also arrive along the axons of the cells of the nuclei of the thin and cuneate fasciculi, some of which go through the lower cerebellar peduncles to its vermis.
In addition, the axons of the cells of the vestibular nuclei go to the cerebellum as part of the inferior peduncle - mainly from the vestibular lateral nucleus of Deiters, they end in the nucleus of the cerebellar clivus.
The fibers of the cells of this nucleus, as part of the upper and, possibly, lower cerebellar peduncles, approach the cells of the reticular formation of the brain stem and the vestibular lateral nucleus, from which the conductors form the descending tracts - vestibulospinal and reticular-spinal, ending at the cells of the anterior horns of the spinal cord.
This pathway regulates the balance of the body.
From the cerebellum, through the vestibular lateral nucleus, connections are established with the nuclei of the oculomotor nerves (as part of the medial longitudinal fasciculus).
The cerebellum has numerous connections with almost all lobes of the brain. There are 2 massive beams.
The fronto-pontine-cerebellar is a collection of axons of cells predominantly in the anterior sections of the superior and middle frontal gyri. In the depths of the lobe, they are collected in a compact bundle and form the anterior leg of the internal capsule.
Then the cerebral peduncles pass through the base and end with a synapse at the cells of the pons. The axons of the second neurons move to the opposite side of the pons and, as part of the middle cerebellar peduncle, enter its hemisphere, contacting the cells of the cerebellar cortex. The processes of these neurons approach the dentate nucleus. The fibers of the dentate nucleus cells in the superior cerebellar peduncle reach the red nucleus of the opposite side and carry impulses along the reticular-spinal tract that regulate human posture in an upright position, in particular standing and walking.
Occipitotemporal ponsocerebellar tract - its first neurons are located in the cortex of the occipital and temporal lobes; their axons gather in the subcortical white matter, then, as part of the posterior femur of the internal capsule, they go at the base of the midbrain to the pons nuclei of their side.
The axons of the pontine cells move to the opposite side and reach the cerebellar cortex along the middle peduncle. The fibers of these cells approach the dentate nucleus, which has connections with the brain stem. With the help of these tracts, coordination of the cerebellum with the organs of vision and hearing is ensured.
Ultimately, the existing decussations of the cerebellar afferent and efferent systems lead to homolateral communication of one hemisphere of the cerebellum and the limbs.
When the cerebellar hemisphere is damaged, disorders of its function occur on the same half of the body. Lesions in the lateral funiculus of the spinal cord also cause cerebellar disorders on its half of the body. The cerebral hemispheres are connected to the opposite hemispheres of the cerebellum.
Therefore, when the cerebral hemispheres or the red nucleus are damaged, cerebellar disorders are detected on the opposite side.
History of cell discovery
Schwann - author of cell theory
In the literature devoted to the history of cell theory, a statement has long been made, which is repeated from time to time even today, that the doctrine of cells as structural formations common to plants and animals belongs equally to the botanist M. Schleiden and the zoologist T. Schwann. However, even at the end of the last century, M. Heidenhain, and later F. Studnicka, and especially the Soviet histologist and historian of cell theory Z. S. Katznelson clearly showed that the role of Schleiden and Schwann in the creation of cell theory is unequal. Schwann should be considered the true founder of this theory, who, in addition to the results of his own research, used the observations of Purkin and his students, Schleiden and a number of other botanists and zoologists.
Schwann's cell theory contains three main generalizations - the theory of cell formation, evidence of the cellular structure of all organs and parts of the body, and the extension of these two principles to the growth and development of animals and plants.
The possibility of comparing plant and animal cells and recognizing complete correspondence (homology) between plant and animal cells was a consequence of two provisions from which Schwanl proceeded. He, together with Schleiden, accepted, firstly, that cells are hollow, vesicular formations and, secondly, that in both kingdoms of nature, cells arise from structureless non-cellular matter located inside or between cells; Schwann called the latter cytoblastema. 3. S. Katsnelson expressed a paradoxical and at the same time correct idea that it was these erroneous views on the nature of cells and the method of their origin that allowed Schwann to see their similarities in plants and animals, while a more correct view of animal cells as formations consisting of granular substance and, unlike plant cells, usually devoid of membranes, developed by Purkin, distracted him from the idea of \u200b\u200bthe homology of cells in plants and animals.
Schwann expressed the cell theory as a broad biological generalization in the following words: “The development of the position that for all organic derivatives there is a general principle of formation and that such is cell formation ... can be given the name cell theory.”
THE CONCEPT OF ATAXIA, TYPES OF ATAXIA
Ataxia is a form of erratic movement, taxis - from the Greek. order, a- negation. It occurs when there is a violation of the coordination of the action of muscle groups - agonists (directly carrying out the movement), antagonists (in some phase counteracting the agonists), synergists (helping the work of either agonists or antagonists).
Movements lose coherence, accuracy, smoothness, proportionality and often do not achieve the goal. Such a patient's muscle strength remains sufficient, he has sufficient strength, he has no paresis.
The pathogenetic essence of ataxia consists of:
Violation of reciprocal innervation. The mechanism of reciprocal (coupled) inhibition of spinal motor centers is as follows: the axons of receptor cells (in the spinal ganglia) in the spinal cord are divided into branches, some of them excite motor neurons of the flexor muscles, and others contact with intercalary cells, which have an inhibitory effect on extensor muscle cells.
The complex integrative function of this mechanism also involves cerebellar impulses.
2. Termination of proprioceptive signaling (from muscle spindles, Golgi tendon bodies) along one or another ascending afferent pathway. Information about the degree of muscle tension at any given moment and about the results of the adaptive effects of functional systems ceases to be received.
The side of the motor function that is referred to as reverse afferentation is upset.
Types of ataxia.
1. Sensitive ataxia is associated with simultaneous suffering in coordination of movements and muscle-articular sensation. With severe ataxia in the upper limb, it is difficult to perform the simplest actions. At rest, involuntary movements are sometimes observed in the fingers of the hand, reminiscent of athetosis - pseudoathetosis. Coordination of movements is also impaired in the lower extremities, which is confirmed by missed hits and jerking movements when performing the heel-knee test.
Muscle tone in the affected limbs is reduced in both the flexor and extensor muscles. In a standing position, staggering is noted, especially when performing the Romberg test.
Movement becomes uncertain, the patient walks with his head down, controlling the act of walking with the help of vision. Thus, sensitive ataxia is always combined with a disorder of deep sensitivity and functional separation of individual segments of the limbs from the higher areas of the brain. Another characteristic feature of this type of ataxia is that it intensifies when the control of the visual analyzer is turned off.
Sensitive ataxia with damage to the posterior cords of the lower half of the spinal cord (for example, with syphilis, funicular myelosis - B12) may be accompanied by the disappearance of deep reflexes in the lower extremities, which is explained by degeneration of the collaterals of the thin bundle fibers, which are the afferent part of the deep reflex arc.
Cerebellar ataxia.
Associated with damage to the cerebellar systems. Considering that the cerebellar vermis takes part in the regulation of contraction of the muscles of the trunk, and the cerebral cortex of the distal limbs, two forms of cerebellar ataxia are distinguished.
- Static-locomtor - when the cerebellar vermis is damaged, mainly standing and walking are disrupted.
The patient stands with his legs wide apart and sways. When walking, the body deviates to the sides, the gait resembles the gait of a drunk. Turning is especially difficult. Deviations when walking are observed towards the cerebellar lesion. Stability is tested in the Romberg position.
If the cerebellar structures are damaged, the patient in this position sways in the appropriate direction; with staggering in the anteroposterior direction - characteristic of damage to the anterior parts of the cerebellar vermis.
The patient's walking in a straight line is examined, as well as the flank gait—stepping movements to the side. At the same time, pay attention to the clarity of the step and the ability to quickly stop in case of a sudden command. When the cerebellar structures are damaged, the combination of simple movements that make up a sequential chain of complex motor acts is disrupted. This is referred to as asynergy and is determined using the Babinski test.
- Dynamic ataxia - with it, the performance of various voluntary movements of the limbs is impaired.
This type of ataxia mainly depends on damage to the cerebellar hemispheres. This is detected when studying the movements of the upper extremities, for example, when performing the finger-to-toe, heel-knee, test for diadochokinesis, etc.
In addition to disruption of these tests with movements of the limbs, when the cerebellum is damaged, speech is disrupted - as a result of incoordination of the speech motor muscles, speech loses smoothness, becomes explosive, stress is placed on the wrong syllables - scanning speech; The handwriting changes - the handwriting becomes uneven, large - megalography.
Nystagmus is observed - a rhythmic twitching of the eyeballs when looking to the sides or up - a kind of intention tremor of the oculomotor muscles. When the cerebellar structures are damaged, the plane of nystagmus coincides with the direction of voluntary movements of the eyeballs - when looking to the side, the nystagmus is horizontal, when looking up and down - vertical.
Sometimes nystagmus is congenital. Such nystagmus is usually present not only when the eyeballs are abducted to the sides (with tension), but also when looking straight (“spontaneous nystagmus”).
When the cerebellar systems are damaged, muscle tone may change.
The most common symptom is muscle hypotonia: the muscles become flabby, flaccid, and joint hypermobility is possible. This may reduce deep reflexes.
Coordination of movements is impaired when the frontal and temporal lobes and their conductors are damaged. In this case, walking and standing are disrupted, the torso deviates posteriorly and in the direction opposite to the lesion. Missing in the arm and leg (hemiataxia) is detected. With this type of coordination disorder, other signs of damage to the corresponding lobes of the cerebral hemispheres are also found.
Cortical ataxia. Coordination of movements is impaired when the frontal and temporal lobes and their conductors are damaged. In this case, walking and standing are disrupted, the torso deviates posteriorly and in the direction opposite to the lesion. Missing in the arm and leg (hemiataxia) is detected. With this type of coordination disorder, other signs of damage to the corresponding lobes of the cerebral hemispheres are also found.
4. Vestibular ataxia - occurs when the function of the vestibular analyzer, in particular its proprioceptors in the labyrinth, is impaired.
With it, the balance of the body is upset; while walking, the patient deviates towards the affected labyrinth. Systemic dizziness, nausea, and horizontal rotatory nystagmus are characteristic. Hearing may be impaired on the side of the affected labyrinth.
Thus, a disorder of coordination of voluntary movements is observed with damage to both the cerebellum itself and the conductors through which impulses from the muscles, semicircular canals of the inner ear and cerebral cortex are brought to it and taken away from the cerebellum to the motor neurons of the brain stem and spinal cord.
Patients with damage to the cerebellar systems usually do not show any pathological manifestations at rest. Various types of incoordination appear in them only when muscles are tense.
Signs of defeat
Purkinje cells transmit impulses through the neurotransmitters GABA. Gamma-aminobutyric acid is a neurotransmitter that has an inhibitory effect so that a person does not make unnecessary, unnecessary movements. When cerebellar tissue is damaged, the activity of Purkinje cells is disrupted.
As a result, a person often does not lose the ability to move, but rather makes chaotic, meaningless, useless movements. Other signs of damage to the cerebellar structures: impaired fine motor skills, inability to maintain balance, excessively strong contractions of the flexor and extensor muscles.
Purkinje cells form the cortical layer of the cerebellum, where information is stored about all motor skills mastered by a person, about motor programs that are honed and improved.
Cerebellum. Physiology of the basal ganglia
The hemispheres are divided by three grooves into the anterior (old cerebellum), posterior (the youngest formation - Neo cerebellum) and trochlear-modular zone (nodule and flocculus - the most ancient parts of the cerebellum)
From a functional point of view, the cerebellum is usually divided into three parts -
The first is the vestibular cerebellum (nodule, flocculus and parts of the posterior lobe partially adjacent to these formations). Primary signals from the receptors of the vestibular apparatus, as well as secondary sensory signals from the nuclei of the medulla oblongata (vestibular nuclei) approach these structures. Afferent fibers approach the tent nucleus, which is located in the white matter of the tent. The vestibular cerebellum controls eye position, body posture, and gait.
The second functional section of the cerebellum is the spinal cerebellum . It includes the vermis and the areas of the anterior and posterior lobes adjacent to the vermis. It is in this zone that the spinocerebellar tracts end, which transmit information from proprioceptors about the position of the limbs and muscle contractions. This information can arrive to the cerebellum discretely (or continuously). This information is used to regulate movements of the trunk (proximal limbs)
The third is the lateral sections of the cerebellar hemispheres ( cortical cerebellum ). Receives information from the cerebral cortex. These pathways pass through the pontine nuclei and middle cerebellar peduncles. Participates in the regulation of the distal limbs. Participates in planning the sequence of movements and the distribution of phases in movement over time. The cerebellum plays a role in the development of visual and auditory phenomena. Based on this activity, a person can predict by changes in visual phenomena how quickly he is approaching something.
The cerebellum receives information from the inferior olivary nuclei. And the pathways from the vestibular system, spinal cord and cerebral cortex approach the inferior olives. The afferent olivocerebellar tract to the cerebellum begins from the inferior olives. This tract crosses in the midline and enters the cerebellum and the fibers of this tract belong to the so-called climbing fibers. Climbing fibers transmit excitation to the cerebellar nuclei and also activate the main cells of the cerebellar cortex - Purkinje cells . All other afferent pathways to the cerebellum consist of mossy fibers. Mossy fibers have an excitatory effect on the cerebellar nuclei and activate granule cells . The cerebellum receives afferent information from:
Spinal cord , from proprioceptors of muscles, tendons, joints along the ventral and dorsal spinocerebellar tracts. The second source is the vestibular nuclei . Third , information comes from the cerebral cortex, which contains copies of motor commands that the cortex sends to the spinal cord to execute movements. The fourth source is the reticular formation from which diffuse information flows to the neurons of the cerebellar cortex. The cerebellum also receives impulses from visual and auditory receptors, from the superior and inferior colliculi.
The efferent pathways of the cerebellum begin from its 4 nuclei - the dentate, spherical, cortical and tent nucleus. From the cerebellar nuclei, the impulse is sent to the motor centers - the red nucleus, the vestibular nucleus, and the nuclei of the reticular formation. And also from the cerebellum, efferent pathways through the ventrolateral part of the thalamus opticum, information is transmitted to the motor and somatosensory areas of the cerebral cortex. The main cells that provide the output signal from the cerebellum are Purkinje cells - large inhibitory neurons. All output signals are braking in nature. In the cerebellar cortex there are 5 types of cells - Purkinje cells (a very developed dendritic tree). Purkinje cells - 15,000,000 in the cerebellar cortex, Golgi cells, basket-shaped, granular, stellate. The cells together with their fibers are the cerebellar cortex. The cerebellar cortex makes up 10% of the cerebral cortex (by mass). And the area of the cerebellar cortex is 75% of the cerebral cortex - due to numerous folds. There are three layers: superficial - molecular, middle - Purkinje cells, internal - granular.
The white matter contains the cerebellar nuclei. Information goes to the cerebellum on 2 types of fibers - climbing - Purkinje cells, mossy - granule cells. Granular cells have a peculiarity - their axon goes from the granular to the surface layer, where it is T-shapedly divided into parallel fibers. These fibers from granule cells form excitatory synapses on 4 cells of the cerebellum. They have a weaker excitatory effect than climbing fibers on Purkinje cells. 4 of these cell types are inhibitory. Basket and stellate cells inhibit Purkinje cells. Golgi cells inhibit grain cells. At the beginning, afferent fibers excite the cerebellar nuclei i.e. the first signal from the cerebellar nuclei will be excitatory, but later, when the Purkinje cell is excited, it will already have an inhibitory effect on the cerebellar nuclei. At the beginning of movement, the cerebellum amplifies the motor signal.
All our movements are pendulum-like; during movement, inertia appears. When we strive to achieve some goal, the hand “passes” this goal, then the cortex gives a signal and everything starts again. To prevent this from happening, the cerebellum turns on and off the antagonist muscles in time. During the influence of the cerebellum, smoothness is achieved. Purkinje cells store information necessary to coordinate movements. The impulse from the foot to the cortex reaches in 0.25 ms. Information from proprioceptors does not give a real state - it shows speed. This information is used by the brain to plan the next stage of movement. Complex work takes place to coordinate movements. Planning of the visual image occurs - the cortex, based on work with the cerebellum, predicts what will happen next.
The cerebellum is a comparison apparatus. It receives information from muscle proprioceptors and stores commands for movement. It analyzes information and commands. The cerebellum can make corrections. Feedback from visual and auditory analyzers helps us with this. You can enter information only when the movements are performed slowly. Fast movements - throwing a ball into a hoop, using musical instruments. High speed – ballistic movements. Speech is also a ballistic movement. The program is formed by the interaction of the cerebellum, the cerebral hemispheres during the learning of movement, and then stored in the cerebellum and the cortex, when necessary, retrieves the necessary information. Purkinje cells learn. When they are already trained, the movements are coordinated.
When it is damaged, various symptoms occur.
Removal of the cerebellum. With damage to the cerebellum - Stage of loss of function, stage of compensation
- Ataxia is the inability to perform a sequence of movements (drunk gait - staggering, legs wide apart, which especially affects turns).
- Astasia - muscles lose the ability to perform tetanic contraction. Therefore, when trying to contract, a trembling occurs. Cerebellar tremor. At rest, when a person does not attempt to move, there is no trembling.
- Intention tremor - trembling occurs when trying to make a movement
- Distance is a violation of muscle tone. First atony, then hypertension
- Asthenia – easy fatigue.
- Adiadochokinesis - the inability to perform opposite movements - supposition, pronation.
- Dysmetria is a violation of the ability to judge distances and the appearance of overshooting.
- Asynergia – is expressed in the fact that movements cease to be smooth, become jerky, the relationship is disrupted
- Disequilibration is a violation of balance.
Abasia – when the body is disturbed in space. The cerebellum also controls autonomic reactions. With cerebellar disorders, disturbances in heart contraction, changes in blood pressure, and changes in muscle tone in the intestines are observed. Regulation of autonomic functions is carried out through the reticular formation and the hypothalamic region.
Physiology of the basal ganglia.
The basal ganglia include a complex of neuronal nodes of the gray matter, which are located in the white matter of the cerebral hemispheres. These formations are called the striopolitan system. It includes the caudate nucleus and putamen - together they form the striatum . The globus pallidus in section consists of 2 segments - outer and inner. The outer segment of the globus pallidus has a common origin with the striatum. The internal segment develops from the gray matter of the diencephalon. These formations have a close connection with the subthalamic nuclei of the diencephalon, with the substantia nigra of the midbrain, which consists of two parts - the ventral part (reticular) and dorsal (compact).
Pars compacta neurons produce dopamine. And the reticular part of the substantia nigra in structure and function resembles the neurons of the inner segment of the globus pallidus.
The substantia nigra forms connections with the anterior ventral nucleus of the visual thalamus, the colliculus colliculi, the pontine nuclei, and bilateral connections with the striatum. These formations receive afferent signals and themselves form efferent pathways. Sensory pathways to the basal ganglia come from the cerebral cortex and the main afferent pathway begins from the motor and premotor cortex.
Cortical areas 2,4,6,8. These pathways go to the striatum and globus pallidus. There is a certain topography of the projection of the muscles of the dorsal part of the shell - the muscles of the legs, arms, and in the ventral part - the mouth and face. From the segments of the globus pallidus there are paths to the visual thalamus, the anterior ventral and ventrolateral nuclei, from which information will be returned to the cortex.
The pathways to the basal ganglia from the visual thalamus are of great importance. Provide sensory information. Influences from the cerebellum are also transmitted to the basal ganglia through the optic thalamus. There are also sensory pathways to the striatum from the substantia nigra . Efferent pathways are represented by connections of the striatum with the globus pallidus, with the substantia nigra, reticular formation of the brain stem; from the globus pallidus there are paths to the red nucleus, to the subthalamic nuclei, to the nuclei of the hypothalamus and visual thalamus. At the subcortical level there are complex circular interactions.
Connections between the cerebral cortex, the thalamus opticus, the basal ganglia and again the cortex form two pathways: direct (facilitates the passage of impulses) and indirect (inhibitory)
Indirect path. Has an inhibitory effect. This inhibitory pathway goes from the striatum to the outer segment of the globus pallidus and the striatum inhibits the outer segment of the globus pallidus. The outer segment of the globus pallidus inhibits the body of Lewis, which normally has an exciting effect on the internal segment of the globus pallidus. In this chain there are two sequential braking.
Substantia nigra (produces dopamine) In the striatum there are 2 types of receptors D1 - excitatory, D2 - inhibitory. The striatum with the substantia nigra has two inhibitory pathways. The substantia nigra inhibits the striatum with dopamine, and the striatum inhibits the substantia nigra with GABA. High copper content in the substantia nigra, the blue spot of the brain stem. The emergence of the striopolitan system was necessary for the movement of the body in space - swimming, crawling, flying. This system forms a connection with the subcortical motor nuclei (red nucleus, tegmentum of the midbrain, nuclei of the reticular formation, vestibular nuclei) From these formations there are descending pathways to the spinal cord. All this together forms the extrapyramidal system.
Motor activity is realized through the pyramidal system - descending pathways. Each hemisphere is connected to the opposite half of the body. In the spinal cord with alpha motor neurons. All our desires are realized through the pyramid system. It works with the cerebellum, the extrapyramidal system and builds several circuits - the cerebellar cortex, the cortex, the extrapyramidal system. The origin of thought arises in the cortex. In order to accomplish it, you need a movement plan. Which includes several components. They are connected into one image. For this you need programs. Programs for fast movements - in the cerebellum . Slow ones - in the basal ganglia. Cora selects the necessary programs. It creates a single general program that will be implemented through the spinal pathways. To throw the ball into the hoop, we need to take a certain position, distribute muscle tone - this is all on a subconscious level - the extrapyramidal system. When everything is ready, the movement itself will take place. The striopolitan system can provide stereotypical learned movements - walking, swimming, cycling, but only when they are learned. When performing a movement, the striopolitan system determines the scale of movements - the amplitude of movements. The scale is determined by the striopolitar system. Hypotonia - decreased tone with hyperkinesis - increased motor activity.
Symptoms of basal ganglia damage
Pure hyperkinesis (accompanied by a decrease in muscle tone) includes
- Chorea - associated with degenerative lesions of the caudate nucleus and manifests itself in the occurrence of fast dancing movements. Rich facial expressions appear, continuous play with fingers, smacking, and develops as a result of rheumatic lesions. All movements are involuntary
-Athetosis - caused by damage to the putamen and globus pallidus and is characterized by slow, wriggling movements - worm-like movements that begin from the distal parts of the limbs and gradually move to the proximal ones.
-Ballism - sweeping movements of the upper and lower limbs
-Huntington's disease is the loss of cholinergic and GABA-secreting neurons of the striatum. This is a genetic disease. It develops as a result of the appearance of an abnormal gene on chromosome 4. Develops from 14 to 50 years, accompanied by movements characteristic of “Chorea” and at the same time progressive dementia develops. The disease leads to death after 15-20 years.
Hyperkinesis in combination with hypertension - Parkinson's disease (decreased dopamine production in the neurons of the compact part of the substantia nigra. The substantia nigra has an inhibitory effect on the striatum. Thus, the content of dopamine in the striatum decreases. Symptoms - a decrease in dopamine to 50% of normal. At the same time, the content decreases and norepinephrine in the hypothalamus.). Symptoms - small movements of the fingers, facial expressions, hypertension (increased muscle tone, mainly flexors. Posture - arms brought to the body, knees bent, head pressed. Trembling at rest - Trenor , mask-like face, slow speech). The symptom of a folding knife is an attempt to bend the arm at the elbow joint - at first there is great resistance, and then easily. The cogwheel symptom is a periodic change of increasing and decreasing tone.
Eldof drugs are administered - they can penetrate the blood-brain barrier and are converted into dopamine. Blockers that destroy norepinephrine and dopamine help. There are attempts to implant cells taken from dead newborns from the substantia nigra
First descriptions of cells
The idea of the discreteness of animal and plant organisms, i.e., of their construction from separate units, sometimes called “cells” (R. Hooke), sometimes “bags” or “bubbles” (M. Malpighi, N. Grew), sometimes “grains” "(K. Wolf), for a long time remained devoid of specific content, since nothing was known about the nature of these formations. The descriptions of F. Fontana (1781), who saw and depicted nuclei and even nucleoli in eel skin cells, went unnoticed; Fontana, of course, was far from understanding the meaning and significance of his observations. Even at the beginning of the 19th century. abstract views were extended to the microscopic structure of organized bodies. For example, in the “Textbook of Natural Philosophy” (1809) by L. Oken, living bodies were described as accumulations of particles, which he called “organic crystals”, “mucous vesicles”, “organic points”, “galvanic vesicles” and even “ciliates”.
The invention of the achromatic microscope and the constant improvement of its optical capabilities made it possible to approach the study of the true structure of cells, primarily plant ones; At first, we were able to see the most noticeable structural formation in them—the shell. It became possible to talk about the true discreteness of the body of higher plants only after in 1812 the German botanist Moldenhauer managed to separate their constituent cells from each other using the method of maceration.